

Revealing past memories: proactive interference and ketamine-induced memory deficits
- PMID: 18434529
- PMCID: PMC6670951
- DOI: 10.1523/JNEUROSCI.0742-07.2008
Revealing past memories: proactive interference and ketamine-induced memory deficits
Abstract
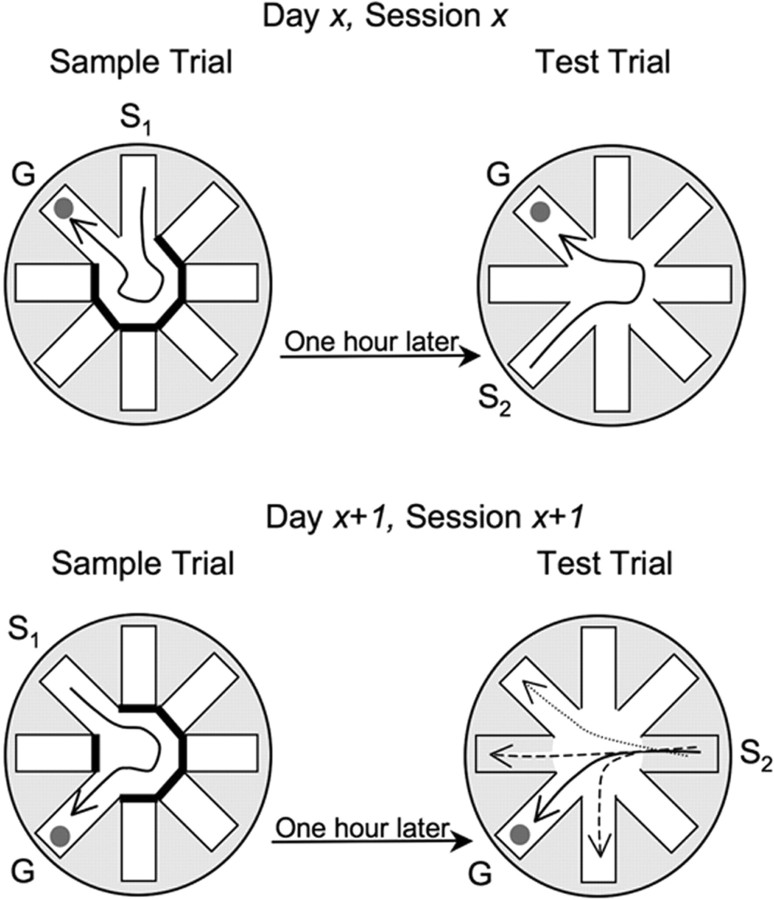
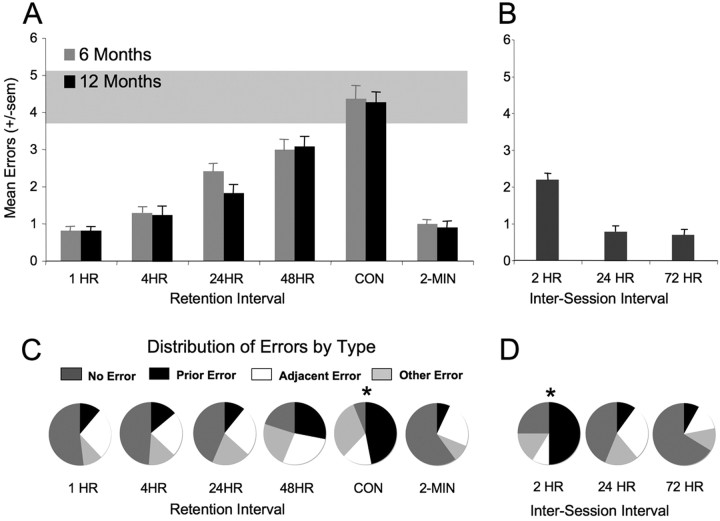
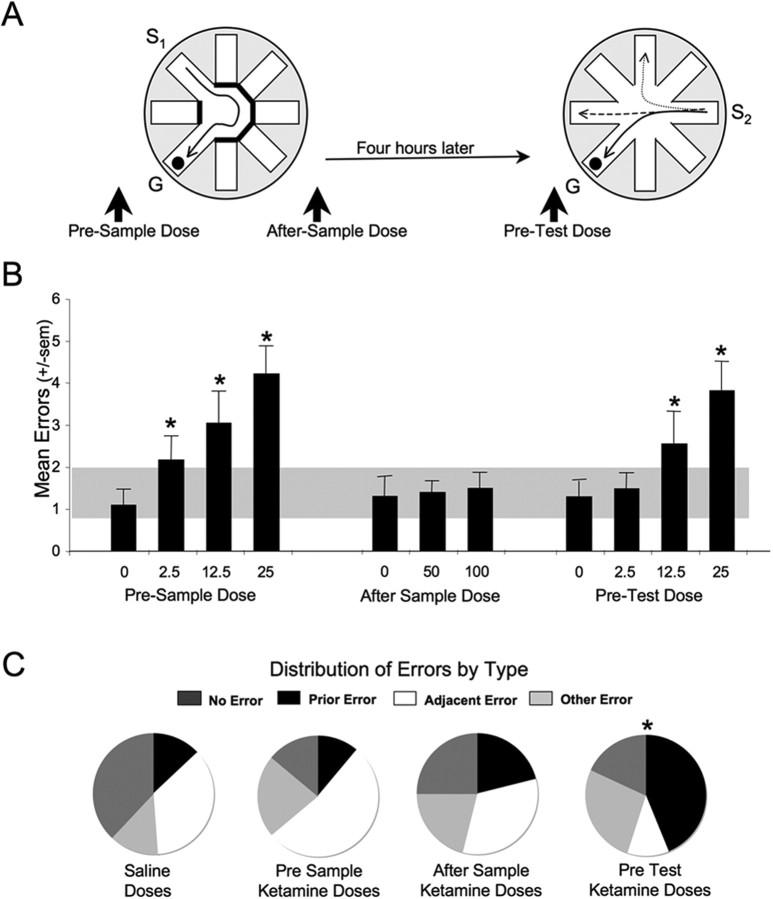
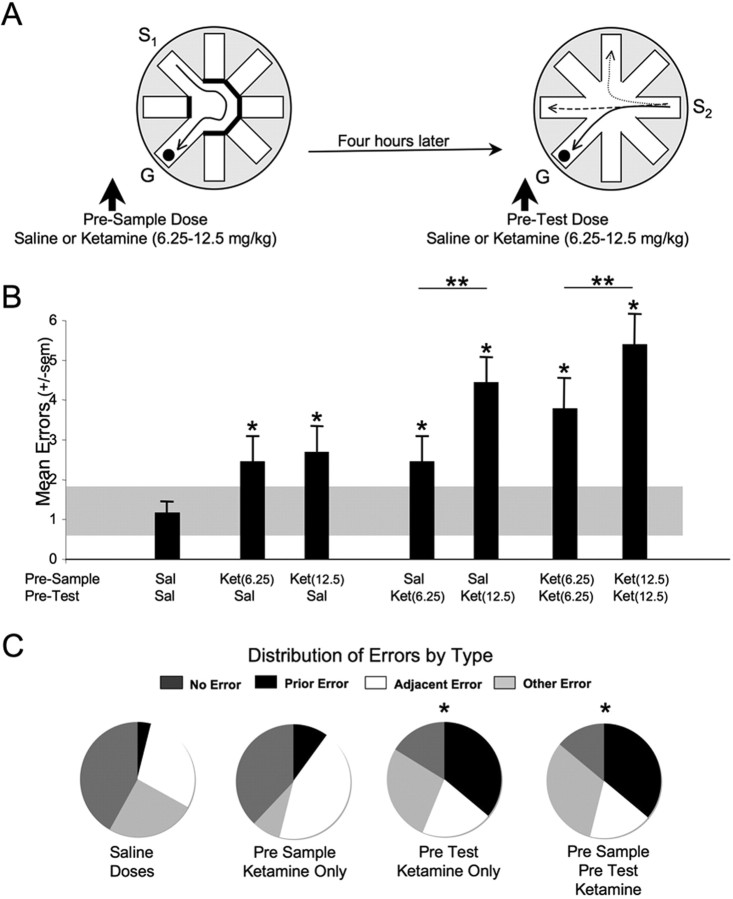
Memories of events that occur often are sensitive to interference from memories of similar events. Proactive interference plays an important and often unexamined role in memory testing for spatially and temporally unique events ("episodes"). Ketamine (NMDA receptor antagonist) treatment in humans and other mammals induces a constellation of cognitive deficits, including impairments in working and episodic memory. We examined the effects of the ketamine (2.5-100 mg/kg) on the acquisition, retrieval, and retention of memory in a delayed-match-to-place radial water maze task that can be used to assess proactive interference. Ketamine (2.5-25 mg/kg, i.p.) given 20 min before the sample trial, impaired encoding. The first errors made during the test trial were predominantly to arms located spatially adjacent to the goal arm, suggesting an established albeit weakened representation. Ketamine (25-100 mg/kg) given immediately after the sample trial had no effect on retention. Ketamine given before the test trial impaired retrieval. First errors under the influence of ketamine were predominantly to the goal location of the previous session. Thus, ketamine treatment promoted proactive interference. These memory deficits were not state dependent, because ketamine treatment at both encoding and retrieval only increased the number of errors during the test session. These data demonstrate the competing influence of distinct memory representations during the performance of a memory task in the rat. Furthermore, they demonstrate the subtle disruptive effects of the NMDA antagonist ketamine on both encoding and retrieval. Specifically, ketamine treatment disrupted retrieval by promoting proactive interference from previous episodic representations.
Figures
References
-
- Bannerman DM, Rawlins JN, Good MA. The drugs don't work—or do they? Pharmacological and transgenic studies of the contribution of NMDA and GluR-A-containing AMPA receptors to hippocampal-dependent memory. Psychopharmacology. 2006;188:552–566. - PubMed
-
- Caramanos Z, Shapiro ML. Spatial memory and N-methyl-d-aspartate receptor antagonists APV and MK-801: memory impairments depend n familiarity with the environment, drug dose, and training duration. Behav Neurosci. 1994;108:30–43. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous