

n-Alcohols inhibit voltage-gated Na+ channels expressed in Xenopus oocytes
- PMID: 18434586
- PMCID: PMC2575017
- DOI: 10.1124/jpet.108.138370
n-Alcohols inhibit voltage-gated Na+ channels expressed in Xenopus oocytes
Abstract
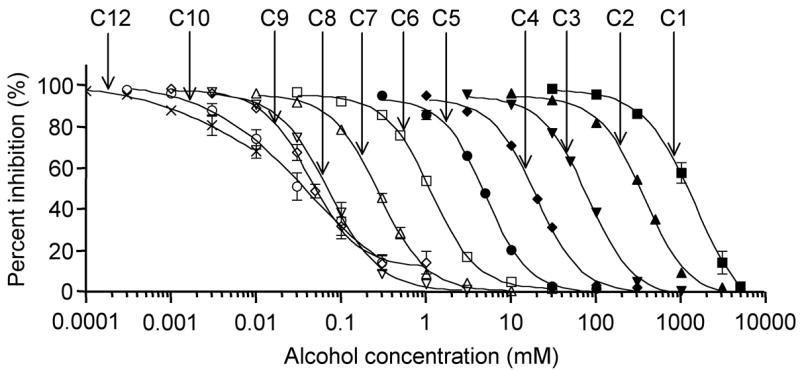
Voltage-gated sodium channels are essential for the initiation and propagation of action potentials in excitable cells and are known as a target of local anesthetics. In addition, inhibition of sodium channels by volatile anesthetics has been proposed as a mechanism of general anesthesia. The n-alcohols produce anesthesia, and their potency increases with carbon number until a "cut-off" is reached. In this study, we examined effects of a range of n-alcohols on Na(v)1.2 subunits to determine the alcohol cut-off for this channel. We also studied the effect of a short-chain alcohol (ethanol) and a long-chain alcohol (octanol) on Na(v)1.2, Na(v)1.4, Na(v)1.6, and Na(v)1.8 subunits, and we investigated the effects of alcohol on channel kinetics. Ethanol and octanol inhibited sodium currents of all subunits, and the inhibition of the Na(v)1.2 channel by n-alcohols indicated a cut-off at nonanol. Ethanol and octanol produced open-channel block, which was more pronounced for Na(v)1.8 than for the other sodium channels. Inhibition of Na(v)1.2 was due to decreased activation and increased inactivation. These results suggest that sodium channels may have a hydrophobic binding site for n-alcohols and demonstrate the differences in the kinetic mechanisms of inhibition for n-alcohols and inhaled anesthetics.
Figures







References
-
- Catterall WA. From ionic currents to molecular mechanisms: The structure and function of voltage-gated sodium channels. Neuron. 2000;26:13–25. - PubMed
-
- Catterall WA, Goldin AL, Waxman SG. International Union of Pharmacology. XLVII. Nomenclature and structure-function relationships of voltage-gated sodium channels. Pharmacol Rev. 2005;57:397–409. - PubMed
-
- Cordeiro JM, Barajas-Martinez H, Hong K, Burashnikov E, Pfeiffer R, Orsino AM, Wu YS, Hu D, Brugada J, Brugada P, Antzelevitch C, Dumaine R, Brugada R. Compound heterozygous mutations P336L and I1660V in the human cardiac sodium channel associated with the Brugada syndrome. Circulation. 2006;114:2026–2033. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
