

The RING domain of cIAP1 mediates the degradation of RING-bearing inhibitor of apoptosis proteins by distinct pathways
- PMID: 18434593
- PMCID: PMC2441678
- DOI: 10.1091/mbc.e08-01-0107
The RING domain of cIAP1 mediates the degradation of RING-bearing inhibitor of apoptosis proteins by distinct pathways
Abstract
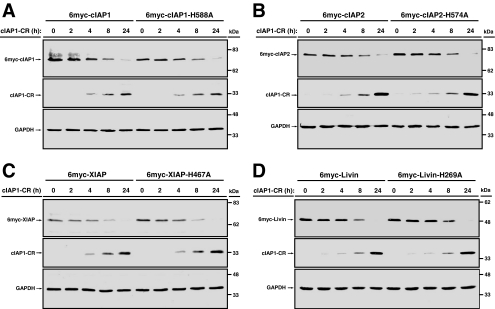
The Inhibitor of Apoptosis proteins (IAPs) are key repressors of apoptosis. Several IAP proteins contain a RING domain that functions as an E3 ubiquitin ligase involved in the ubiquitin-proteasome pathway. Here we investigated the interplay of ubiquitin-proteasome pathway and RING-mediated IAP turnover. We found that the CARD-RING domain of cIAP1 (cIAP1-CR) is capable of down-regulating protein levels of RING-bearing IAPs such as cIAP1, cIAP2, XIAP, and Livin, while sparing NAIP and Survivin, which do not possess a RING domain. To determine whether polyubiquitination was required, we tested the ability of cIAP1-CR to degrade IAPs under conditions that impair ubiquitination modifications. Remarkably, although the ablation of E1 ubiquitin-activating enzyme prevented cIAP1-CR-mediated down-regulation of cIAP1 and cIAP2, there was no impact on degradation of XIAP and Livin. XIAP mutants that were not ubiquitinated in vivo were readily down-regulated by cIAP1-CR. Moreover, XIAP degradation in response to cisplatin and doxorubicin was largely prevented in cIAP1-silenced cells, despite cIAP2 up-regulation. The knockdown of cIAP1 and cIAP2 partially blunted Fas ligand-mediated down-regulation of XIAP and protected cells from cell death. Together, these results show that the E3 ligase RING domain of cIAP1 targets RING-bearing IAPs for proteasomal degradation by ubiquitin-dependent and -independent pathways.
Figures






Similar articles
-
Degradation of survivin by the X-linked inhibitor of apoptosis (XIAP)-XAF1 complex.J Biol Chem. 2007 Sep 7;282(36):26202-9. doi: 10.1074/jbc.M700776200. Epub 2007 Jul 5. J Biol Chem. 2007. PMID: 17613533
-
Molecular determinants of Smac mimetic induced degradation of cIAP1 and cIAP2.Cell Death Differ. 2011 Aug;18(8):1376-86. doi: 10.1038/cdd.2011.10. Epub 2011 Feb 18. Cell Death Differ. 2011. PMID: 21331077 Free PMC article.
-
Smac/DIABLO selectively reduces the levels of c-IAP1 and c-IAP2 but not that of XIAP and livin in HeLa cells.J Biol Chem. 2004 Apr 23;279(17):16963-70. doi: 10.1074/jbc.M401253200. Epub 2004 Feb 11. J Biol Chem. 2004. PMID: 14960576
-
Inhibitors of apoptosis proteins (IAPs) as potential molecular targets for therapy of hematological malignancies.Curr Mol Med. 2011 Nov;11(8):633-49. doi: 10.2174/156652411797536723. Curr Mol Med. 2011. PMID: 21902653 Review.
-
Modulation of immune signalling by inhibitors of apoptosis.Trends Immunol. 2012 Nov;33(11):535-45. doi: 10.1016/j.it.2012.06.004. Epub 2012 Jul 24. Trends Immunol. 2012. PMID: 22836014 Review.
Cited by
-
Tumor necrosis factor (TNF) signaling, but not TWEAK (TNF-like weak inducer of apoptosis)-triggered cIAP1 (cellular inhibitor of apoptosis protein 1) degradation, requires cIAP1 RING dimerization and E2 binding.J Biol Chem. 2010 Jun 4;285(23):17525-36. doi: 10.1074/jbc.M109.087635. Epub 2010 Mar 30. J Biol Chem. 2010. PMID: 20356846 Free PMC article.
-
Therapeutic potential and molecular mechanism of a novel, potent, nonpeptide, Smac mimetic SM-164 in combination with TRAIL for cancer treatment.Mol Cancer Ther. 2011 May;10(5):902-14. doi: 10.1158/1535-7163.MCT-10-0864. Epub 2011 Mar 3. Mol Cancer Ther. 2011. PMID: 21372226 Free PMC article.
-
TNFR-associated factor 2 deficiency in B lymphocytes predisposes to chronic lymphocytic leukemia/small lymphocytic lymphoma in mice.J Immunol. 2012 Jul 15;189(2):1053-61. doi: 10.4049/jimmunol.1200814. Epub 2012 Jun 18. J Immunol. 2012. PMID: 22711886 Free PMC article.
-
Distinctive effects of the cellular inhibitor of apoptosis protein c-IAP2 through stabilization by XIAP in glioblastoma multiforme cells.Cell Cycle. 2014;13(6):992-1005. doi: 10.4161/cc.27880. Epub 2014 Jan 22. Cell Cycle. 2014. PMID: 24552816 Free PMC article.
-
Expression of the IAP protein family acts cooperatively to predict prognosis in human bladder cancer patients.Oncol Lett. 2013 Apr;5(4):1278-1284. doi: 10.3892/ol.2013.1150. Epub 2013 Jan 23. Oncol Lett. 2013. PMID: 23599779 Free PMC article.
References
-
- Arora V., Cheung H. H., Plenchette S., Micali O. C., Liston P., Korneluk R. G. Degradation of survivin by the X-linked inhibitor of apoptosis (XIAP)-XAF1 complex. J. Biol. Chem. 2007;282:26202–26209. - PubMed
-
- Chang H., Schimmer A. D. Livin/melanoma inhibitor of apoptosis protein as a potential therapeutic target for the treatment of malignancy. Mol. Cancer Ther. 2007;6:24–30. - PubMed
-
- Cheung H. H., Gurd J. W. Tyrosine phosphorylation of the N-methyl-D-aspartate receptor by exogenous and postsynaptic density-associated Src-family kinases. J. Neurochem. 2001;78:524–534. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
