

Age-related changes in the osteogenic differentiation potential of mouse bone marrow stromal cells
- PMID: 18435580
- PMCID: PMC2679384
- DOI: 10.1359/jbmr.080304
Age-related changes in the osteogenic differentiation potential of mouse bone marrow stromal cells
Abstract
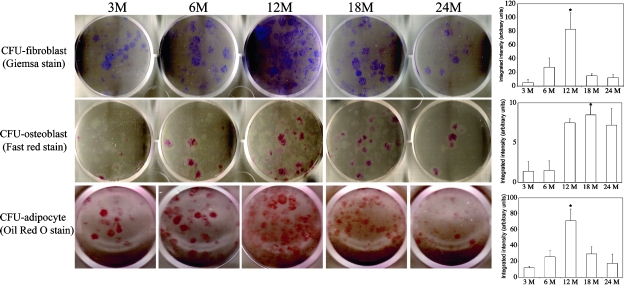
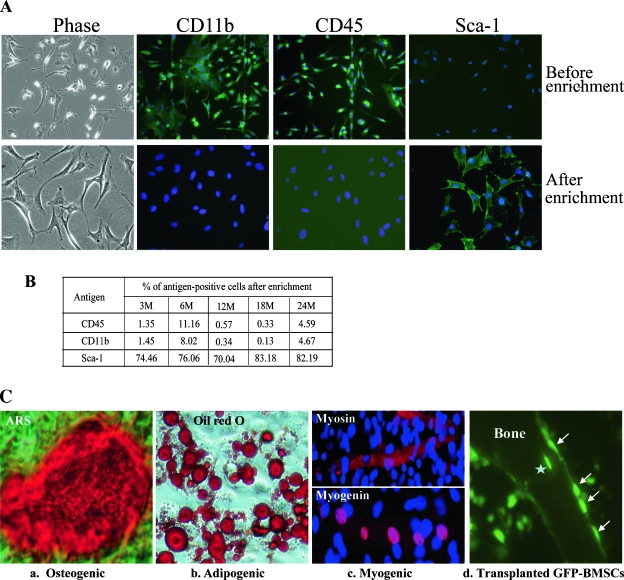
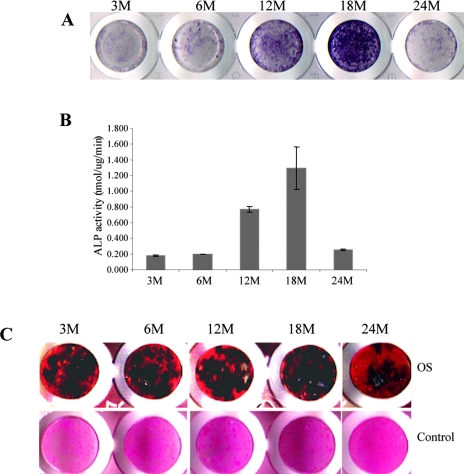
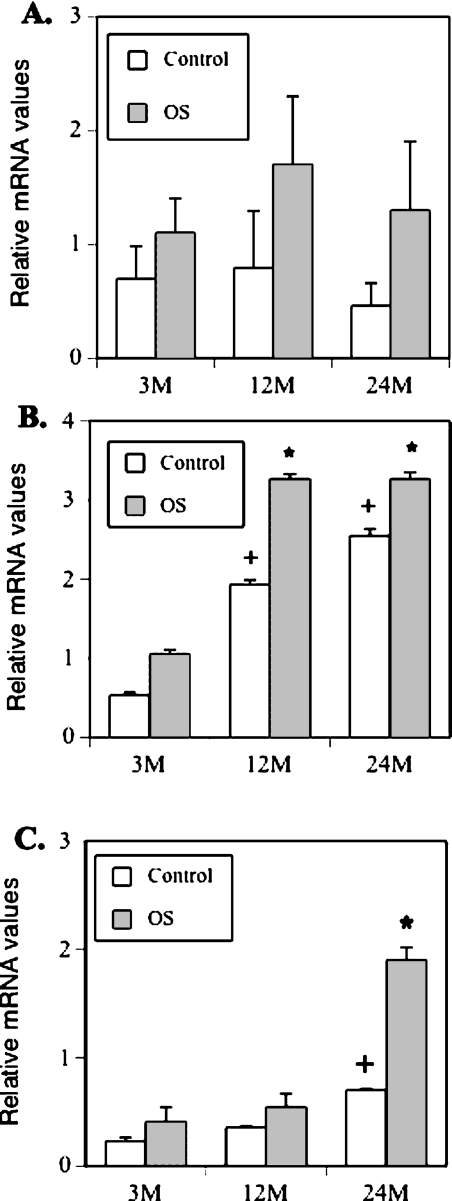
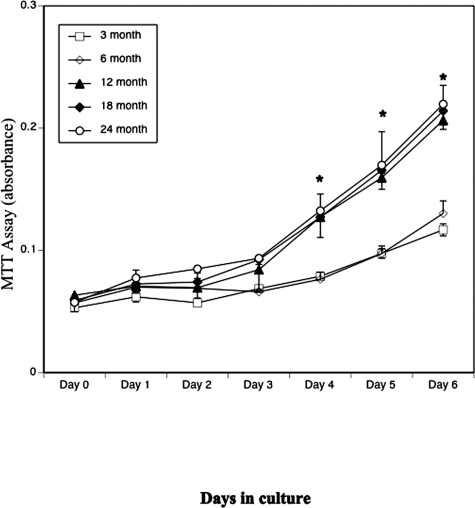
Age-dependent bone loss has been well documented in both human and animal models. Although the underlying causal mechanisms are probably multifactorial, it has been hypothesized that alterations in progenitor cell number or function are important. Little is known regarding the properties of bone marrow stromal cells (BMSCs) or bone progenitor cells during the aging process, so the question of whether aging alters BMSC/progenitor osteogenic differentiation remains unanswered. In this study, we examined age-dependent changes in bone marrow progenitor cell number and differentiation potential between mature (3 and 6 mo old), middle-aged (12 and 18 mo old), and aged (24 mo old) C57BL/6 mice. BMSCs or progenitors were isolated from five age groups of C57BL/6 mice using negative immunodepletion and positive immunoselection approaches. The osteogenic differentiation potential of multipotent BMSCs was determined using standard osteogenic differentiation procedures. Our results show that both BMSC/progenitor number and differentiation potential increase between the ages of 3 and 18 mo and decrease rapidly thereafter with advancing age. These results are consistent with the changes of the mRNA levels of osteoblast lineage-associated genes. Our data suggest that the decline in BMSC number and osteogenic differentiation capacity are important factors contributing to age-related bone loss.
Figures

References
-
- Ferguson VL, Ayers RA, Bateman TA, Simske SJ. Bone development and age-related bone loss in male C57BL/6J mice. Bone. 2003;33:387–398. - PubMed
-
- Gudmundsdottir SL, Indridason OS, Franzson L, Sigurdsson G. Age-related decline in bone mass measured by dual-energy X-ray absorptiometry and quantitative ultrasound in a population-based sample of both sexes: Identification of useful ultrasound thresholds for osteoporosis screening. J Clin Densitom. 2005;8:80–86. - PubMed
-
- Hamrick MW, Shi X, Zhang W, Pennington C, Thakore H, Haque M, Kang B, Isales CM, Fulzele S, Wenger KH. Loss of myostatin (GDF8) function increases osteogenic differentiation of bone marrow-derived mesenchymal stem cells but the osteogenic effect is ablated with unloading. Bone. 2007;40:1544–1553. - PMC - PubMed
-
- Rupprecht M, Pogoda P, Mumme M, Rueger JM, Puschel K, Amling M. Bone microarchitecture of the calcaneus and its changes in aging: A histomorphometric analysis of 60 human specimens. J Orthop Res. 2006;24:664–674. - PubMed
-
- Manolagas SC. Birth and death of bone cells: Basic regulatory mechanisms and implications for the pathogenesis and treatment of osteoporosis. Endocr Rev. 2000;21:115–137. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
