

The Rsu-1-PINCH1-ILK complex is regulated by Ras activation in tumor cells
- PMID: 18436335
- PMCID: PMC2600675
- DOI: 10.1016/j.ejcb.2008.02.011
The Rsu-1-PINCH1-ILK complex is regulated by Ras activation in tumor cells
Abstract
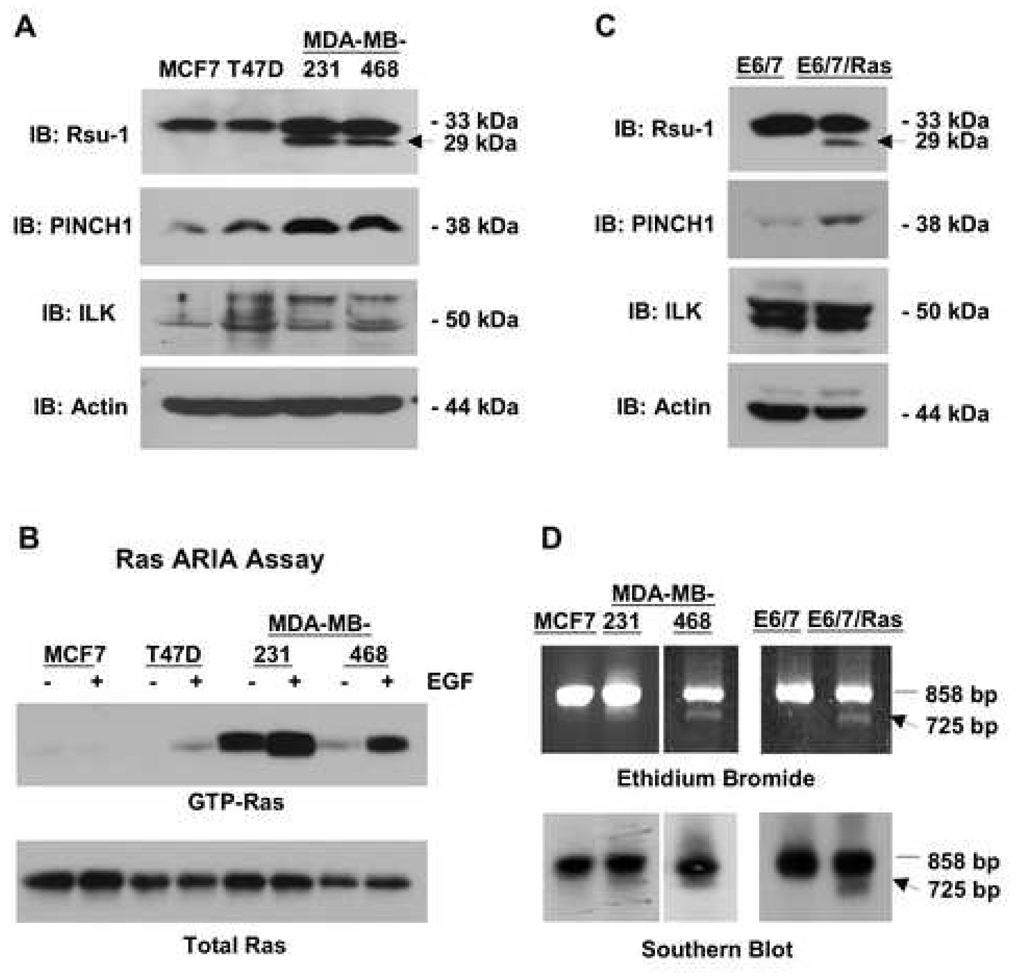
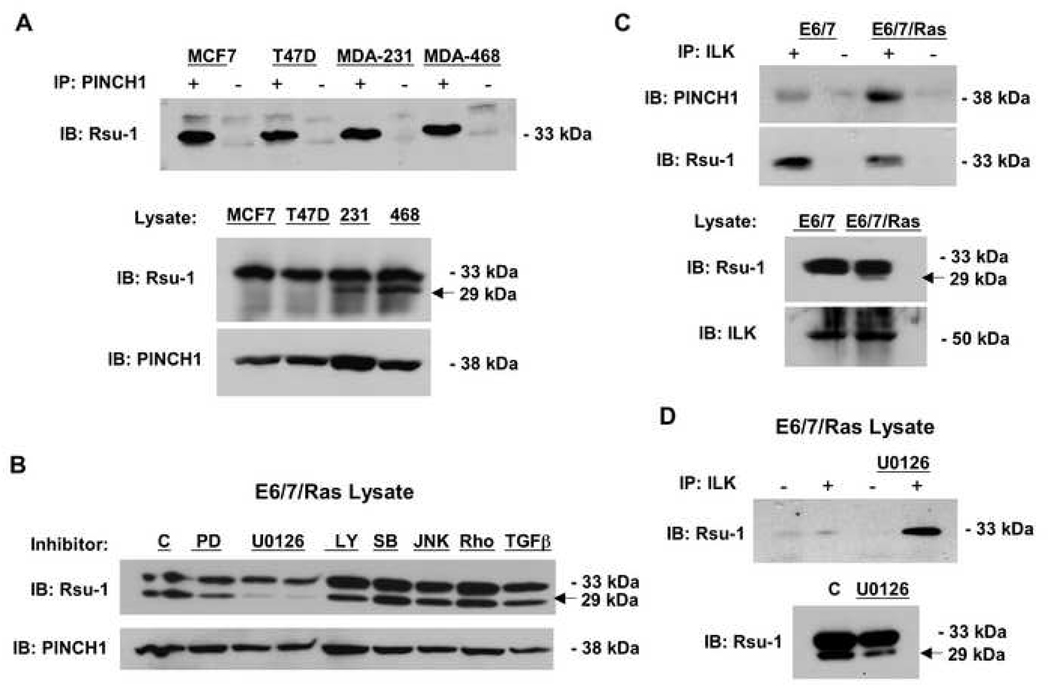
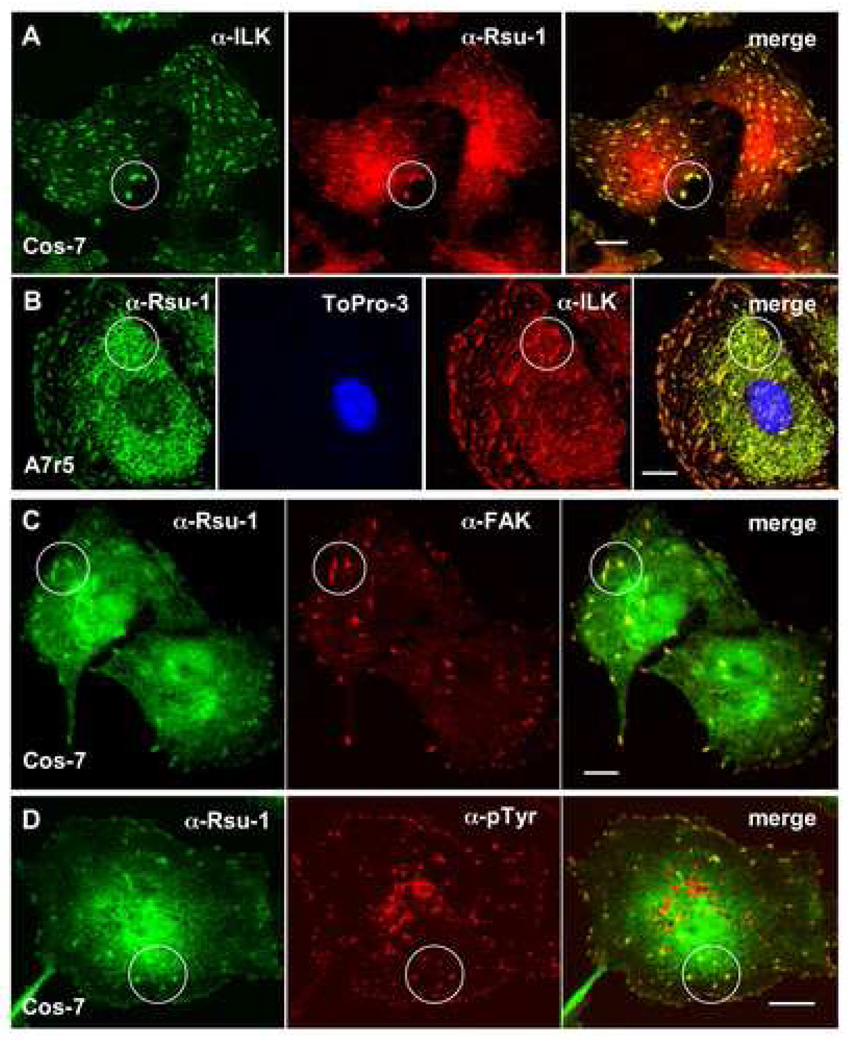
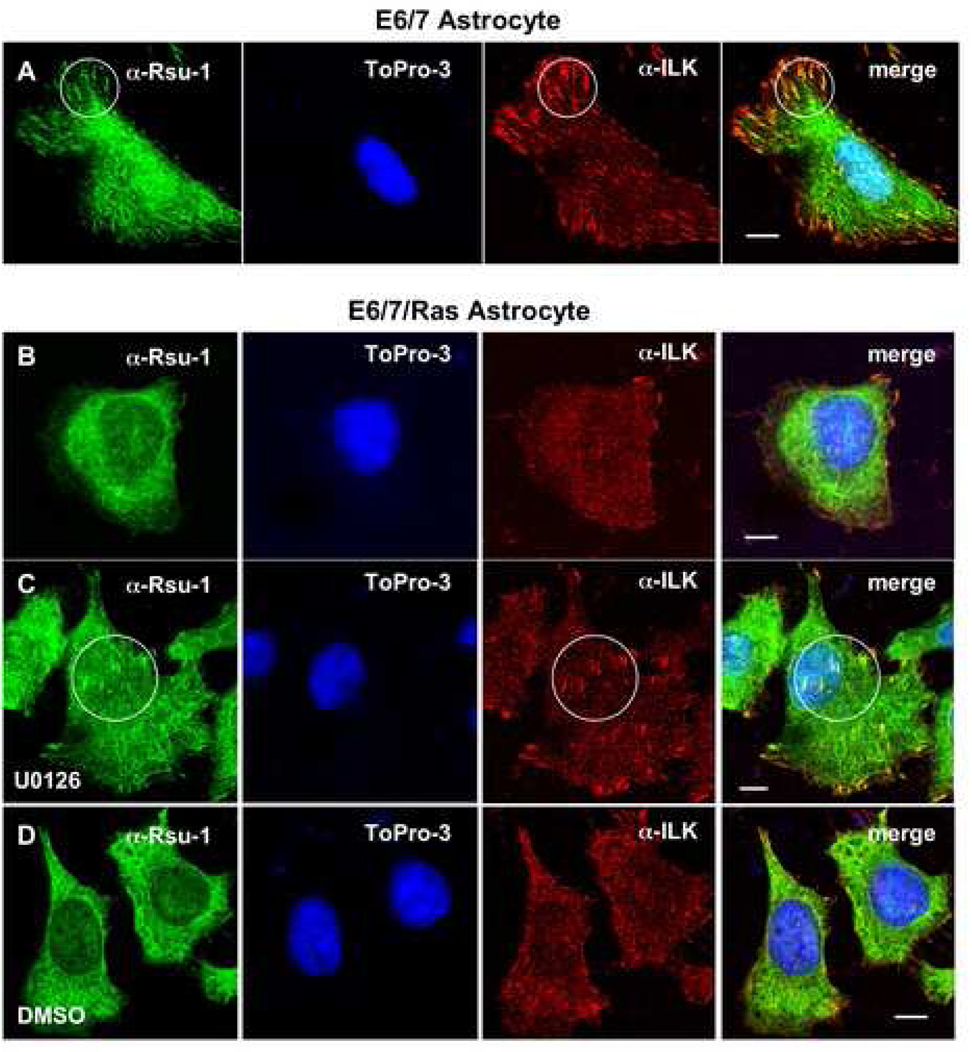
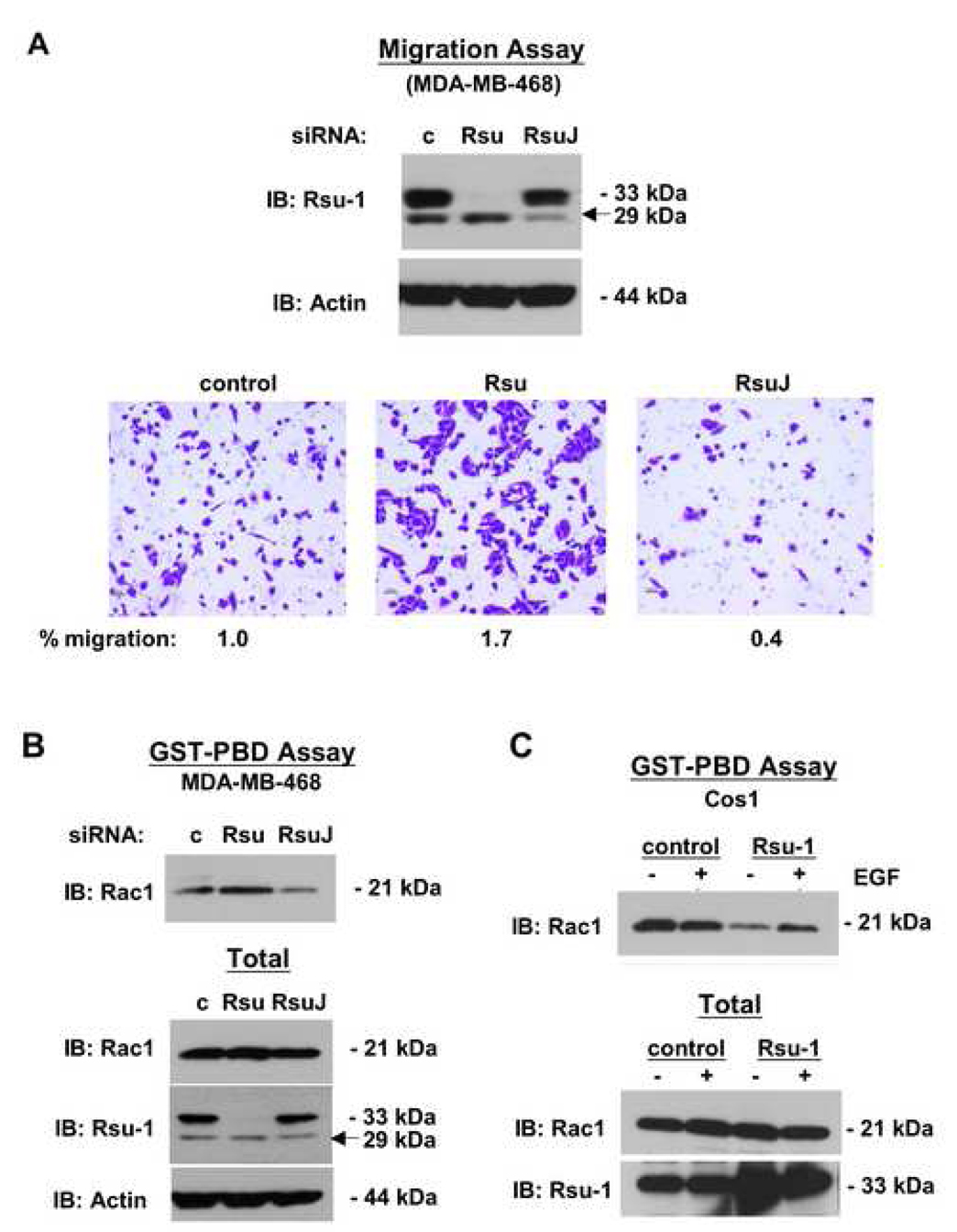
The link between Ras transformation and enhanced cell migration due to altered integrin signaling is well established in tumorigenesis, however there remain gaps in our understanding of its mechanism. The Ras suppressor, Rsu-1, has recently been linked to the IPP (integrin-linked kinase {ILK}, PINCH-1/LIMS1, parvin) focal adhesion complex based on its interaction with the LIM 5 domain of PINCH1. Defining the role of the Rsu1-PINCH1-ILK-parvin complex in tumorigenesis is important because both ILK and PINCH1 are elevated in certain tumors while ectopic expression of Rsu-1 blocks tumorigenesis. Our studies previously identified an alternatively spliced isoform of Rsu-1 in high-grade gliomas. We report here the detection of a truncated (p29) Rsu-1 protein, which correlates with the presence of the alternatively spliced Rsu-1 RNA. This RNA and the respective protein were detected in human tumor cell lines that contain high levels of activated Ras, and inhibitor studies demonstrate that the Mek-ERK pathway regulates expression of this truncated Rsu-1 product. We also show that Rsu-1 co-localizes with ILK at focal contacts and co-immunoprecipitates with the ILK-PINCH1 complex in non-transformed cells, but following Ras transformation the association of Rsu-1 with the PINCH1-ILK complex is greatly reduced. Using a human breast cancer cell line, our in vitro studies demonstrate that the depletion of Rsu-1 full-length protein enhances cell migration coincident with an increase in Rac-GTP while the depletion of the p29 Rsu-1 truncated protein inhibits migration. These findings indicate that Rsu-1 may inhibit cell migration by stabilizing the IPP adhesion complex and that Ras activation perturbs this inhibitory function by modulating both Rsu-1 splicing and association of full-length Rsu-1 with IPP. Hence, our findings demonstrate that Rsu-1 links the Ras pathway with the IPP complex and the perturbations of cell attachment-dependent signaling that occur in the malignant process.
Figures
References
-
- Ahmed N, Riley C, Oliva K, Stutt E, Rice GE, Quinn MA. Integrin-linked kinase expression increases with ovarian tumour grade and is sustained by peritoneal tumour fluid. J. Pathol. 2003;201:229–237. - PubMed
-
- Albini A, Iwamoto Y, Kleinman HK, Martin GR, Aaronson SA, Kozlowski JM, McEwan RN. A rapid in vitro assay for quantitating the invasive potential of tumor cells. Cancer Res. 1987;47:3239–3245. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
