

The acute phase protein haptoglobin regulates host immunity
- PMID: 18436583
- PMCID: PMC3178501
- DOI: 10.1189/jlb.0208100
The acute phase protein haptoglobin regulates host immunity
Abstract
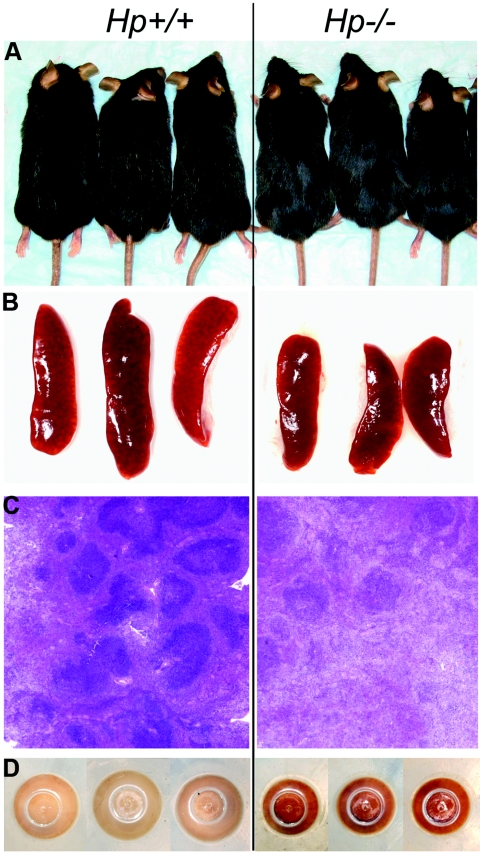
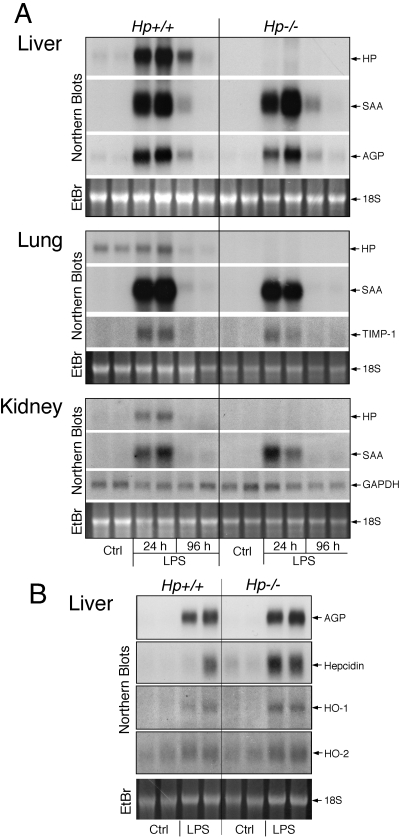
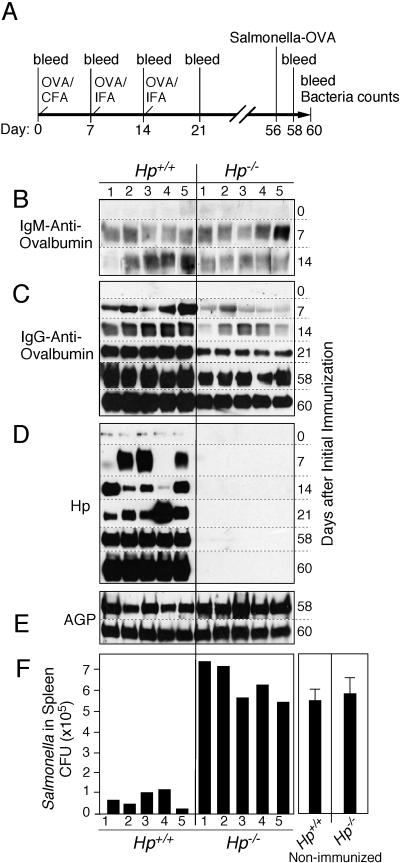
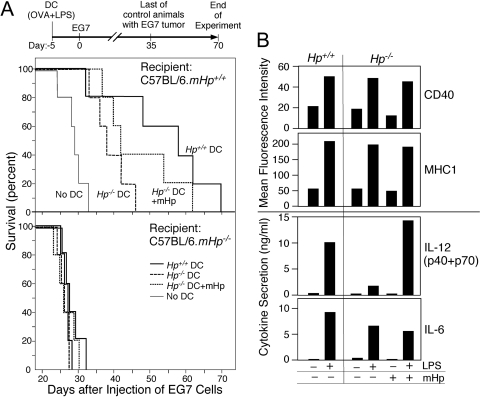
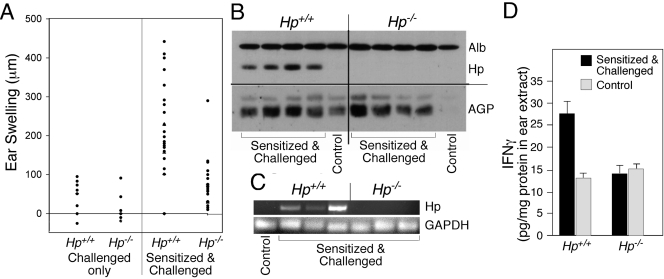
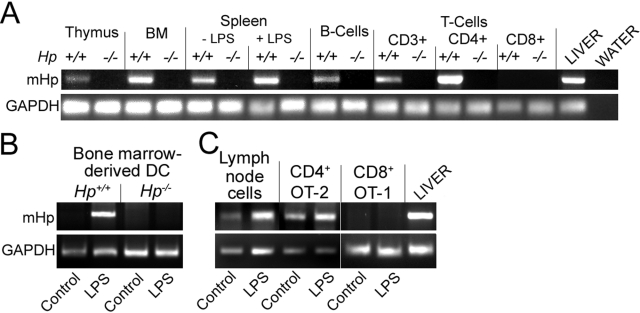
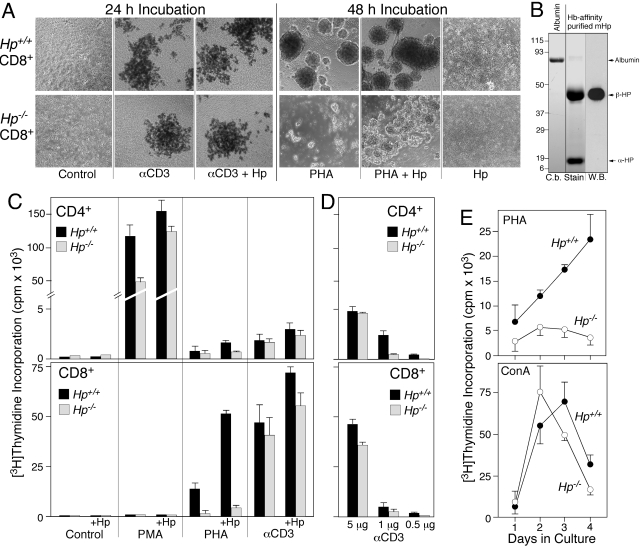
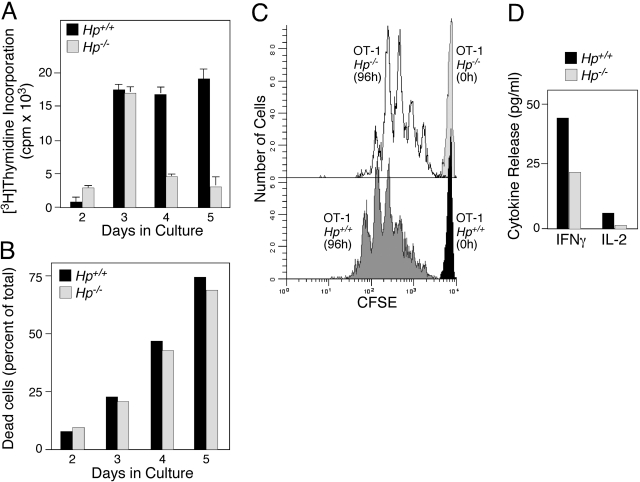
The contribution of acute phase plasma proteins to host immune responses remains poorly characterized. To better understand the role of the acute phase reactant and major hemoglobin-binding protein haptoglobin (Hp) on the function of immune cells, we generated Hp-deficient C57BL/6J mice. These mice exhibit stunted development of lymphoid organs associated with lower counts of mature T and B cells in the blood and secondary lymphoid compartments. Moreover, these mice show markedly reduced adaptive immune responses as represented by reduced accumulation of IgG antibody after immunization with adjuvant and nominal antigen, abrogation of Th1-dominated delayed-type hypersensitivity reaction, loss of mitogenic responses mounted by T cells, and reduced T cell responses conveyed by APCs. Collectively, these defects are in agreement with the observations that Hp-deficient mice are not capable of generating a recall response or deterring a Salmonella infection as well as failing to generate tumor antigen-specific responses. The administration of Hp to lymphocytes in tissue culture partially ameliorates these functional defects, lending further support to our contention that the acute phase response protein Hp has the ability to regulate immune cell responses and host immunity. The phenotype of Hp-deficient mice suggests a major regulatory activity for Hp in supporting proliferation and functional differentiation of B and T cells as part of homeostasis and in response to antigen stimulation.
Figures
References
-
- Jayle M. F., Boussier G., Badin J. Electrophoresis of haptoglobin and of its hemoglobin complex. Bull Soc Chim Biol (Paris) 1952;34:1063–1069. - PubMed
-
- Dobryszycka W. Biological functions of haptoglobin—new pieces to an old puzzle. Eur J Clin Chem Clin Biochem. 1997;35:647–654. - PubMed
-
- Wang Y., Kinzie E., Berger F. G., Lim S. K., Baumann H. Haptoglobin, an inflammation-inducible plasma protein. Redox Rep. 2001;6:379–385. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
