

Neuropilin-1 is a receptor for transforming growth factor beta-1, activates its latent form, and promotes regulatory T cell activity
- PMID: 18436584
- PMCID: PMC2504713
- DOI: 10.1189/jlb.0208090
Neuropilin-1 is a receptor for transforming growth factor beta-1, activates its latent form, and promotes regulatory T cell activity
Abstract
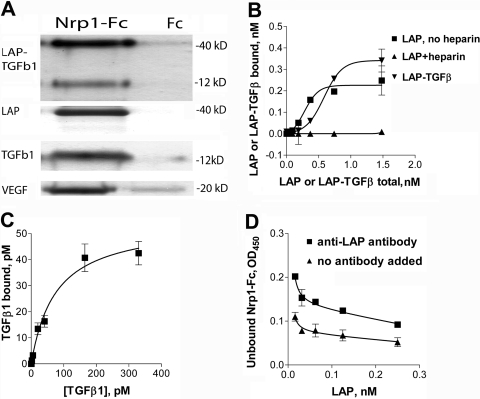
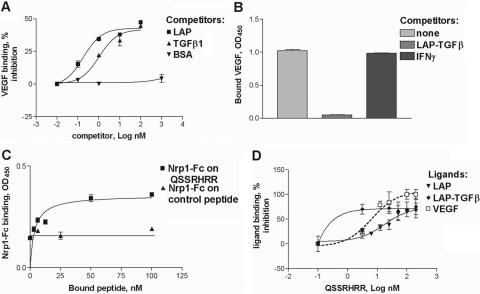
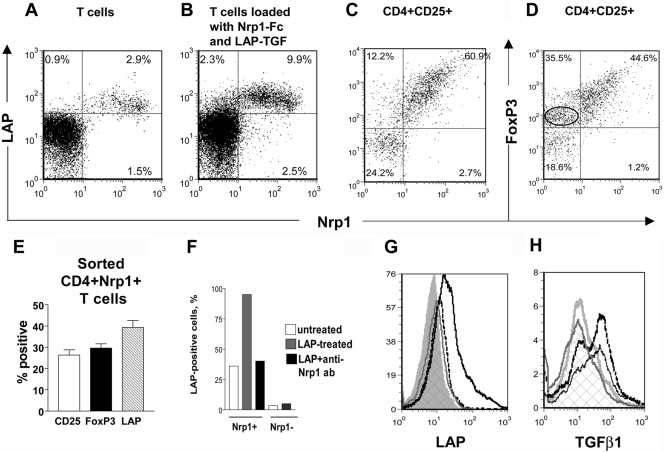
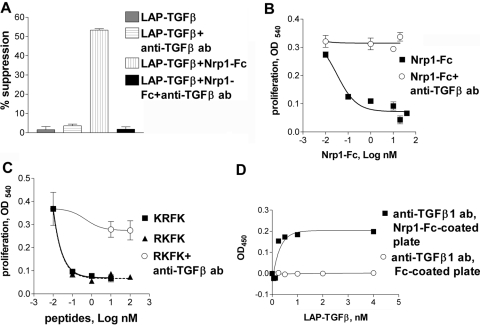
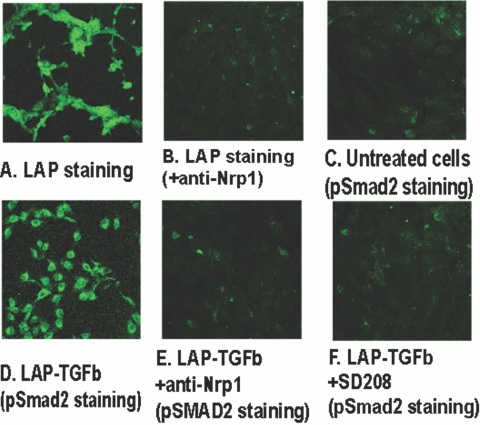
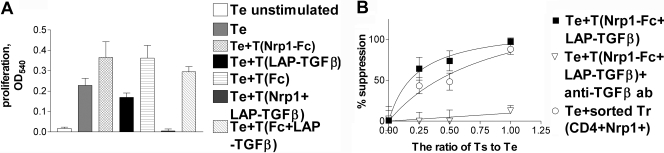
Neuropilin-1 (Nrp1) is a multifunctional protein, identified principally as a receptor for the class 3 semaphorins and members of the vascular endothelial growth factor (VEGF) family, but it is capable of other interactions. It is a marker of regulatory T cells (Tr), which often carry Nrp1 and latency-associated peptide (LAP)-TGF-beta1 (the latent form). The signaling TGF-beta1 receptors bind only active TGF-beta1, and we hypothesized that Nrp1 binds the latent form. Indeed, we found that Nrp1 is a high-affinity receptor for latent and active TGF-beta1. Free LAP, LAP-TGF-beta1, and active TGF-beta1 all competed with VEGF165 for binding to Nrp1. LAP has a basic, arginine-rich C-terminal motif similar to VEGF and peptides that bind to the b1 domain of Nrp1. A C-terminal LAP peptide (QSSRHRR) bound to Nrp1 and inhibited the binding of VEGF and LAP-TGF-beta1. We also analyzed the effects of Nrp1/LAP-TGF-beta1 coexpression on T cell function. Compared with Nrp1(-) cells, sorted Nrp1+ T cells had a much greater capacity to capture LAP-TGF-beta1. Sorted Nrp1(-) T cells captured soluble Nrp1-Fc, and this increased their ability to capture LAP-TGF-beta1. Conventional CD4+CD25(-)Nrp1(-) T cells coated with Nrp1-Fc/LAP-TGF-beta1 acquired strong Tr activity. Moreover, LAP-TGF-beta was activated by Nrp1-Fc and also by a peptide of the b2 domain of Nrp1 (RKFK; similar to a thrombospondin-1 peptide). Breast cancer cells, which express Nrp1, also captured and activated LAP-TGF-beta1 in a Nrp1-dependent manner. Thus, Nrp1 is a receptor for TGF-beta1, activates its latent form, and is relevant to Tr activity and tumor biology.
Figures
References
-
- Staton C A, Kumar I, Reed M W, Brown N. Neuropilins in physiological and pathological angiogenesis. J Pathol. 2007;212:237–248. - PubMed
-
- Roskoski R., Jr Vascular endothelial growth factor (VEGF) signaling in tumor progression. Crit Rev Oncol Hematol. 2007;62:179–213. - PubMed
-
- Lee C C, Kreusch A, McMullan D, Ng K, Spraggon G. Crystal structure of the human neuropilin-1 b1 domain. Structure. 2003;11:99–108. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
