

Mouse Eri1 interacts with the ribosome and catalyzes 5.8S rRNA processing
- PMID: 18438418
- PMCID: PMC3032500
- DOI: 10.1038/nsmb.1417
Mouse Eri1 interacts with the ribosome and catalyzes 5.8S rRNA processing
Abstract
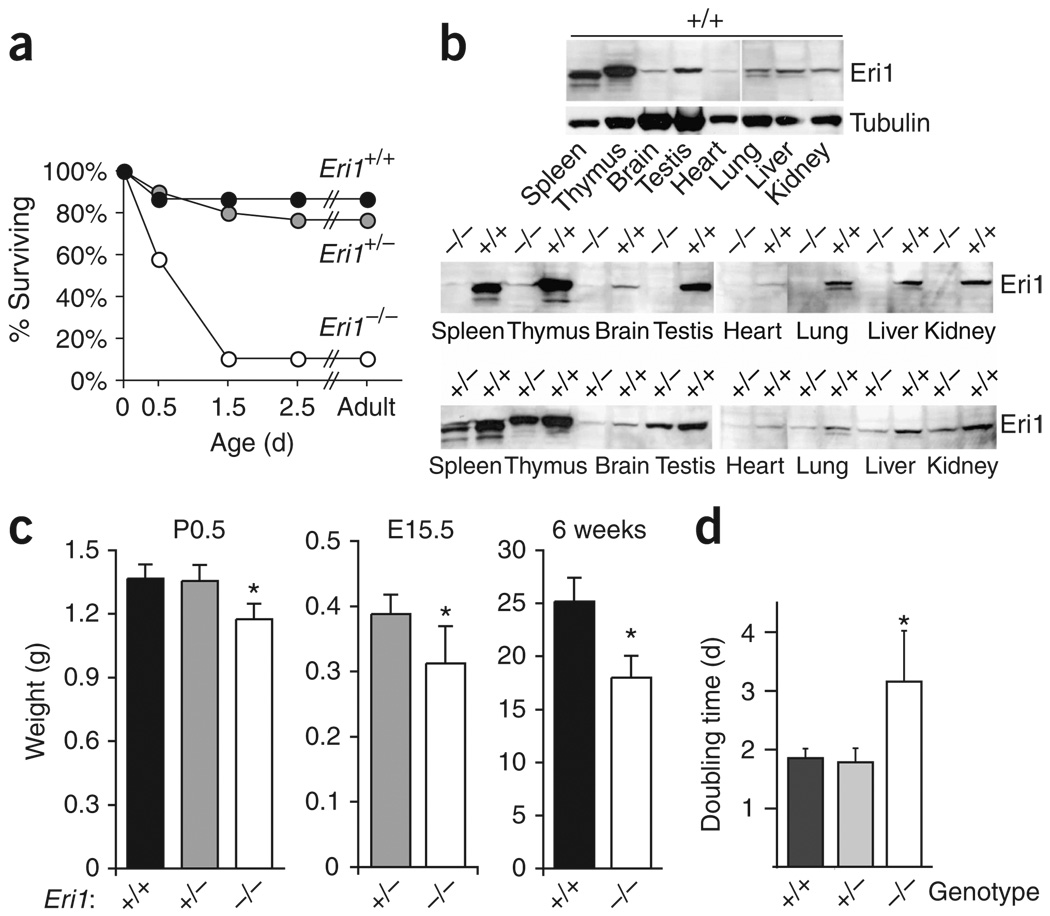
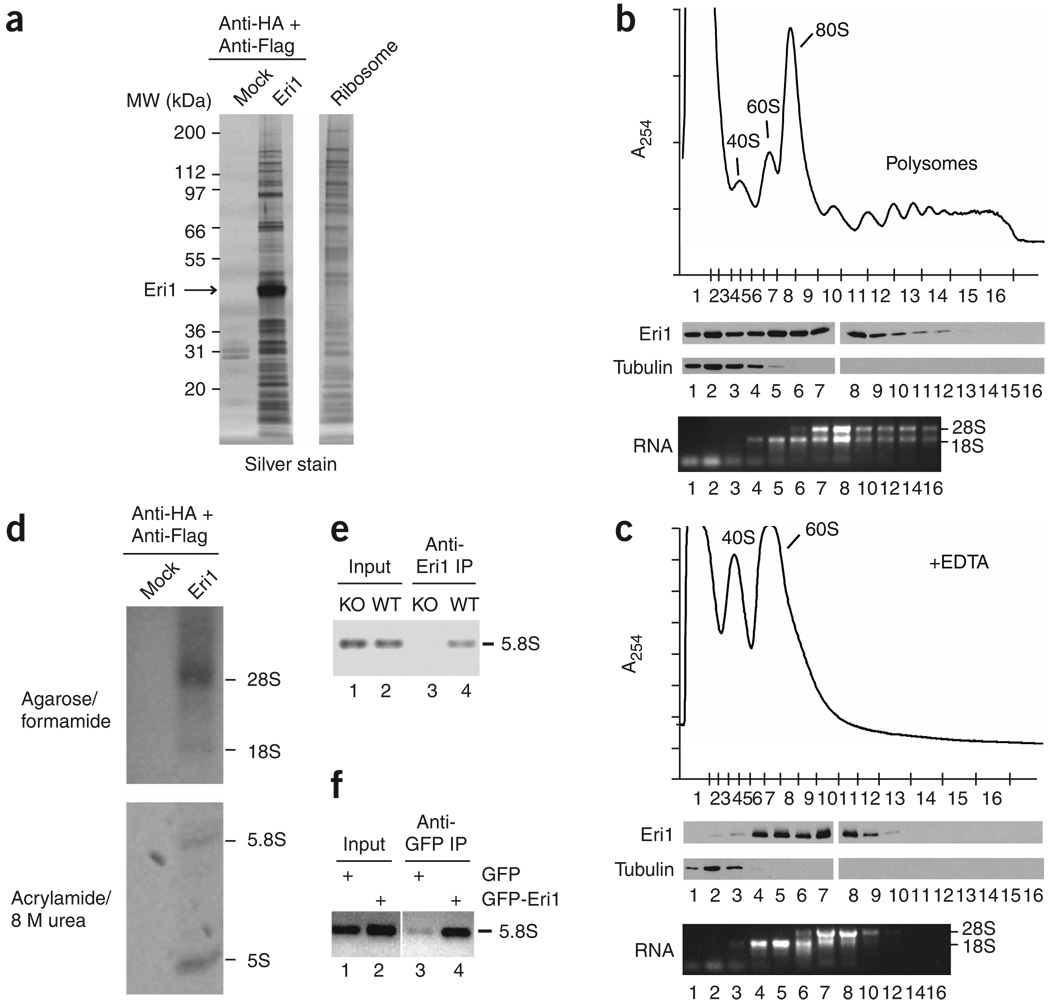
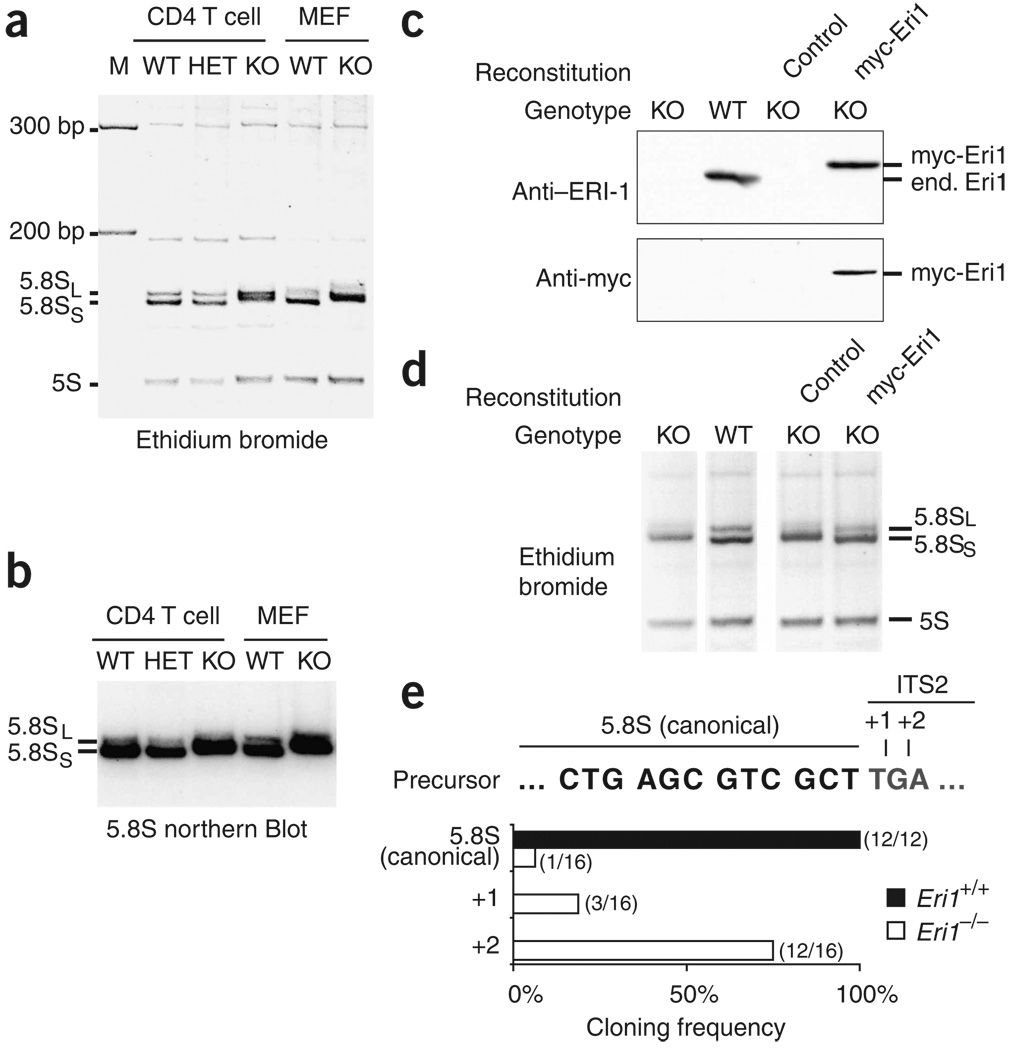
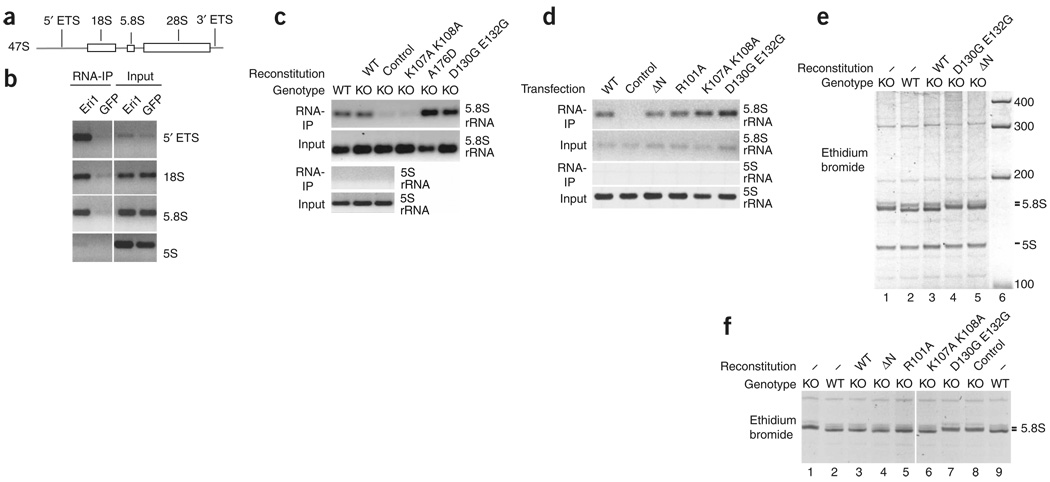
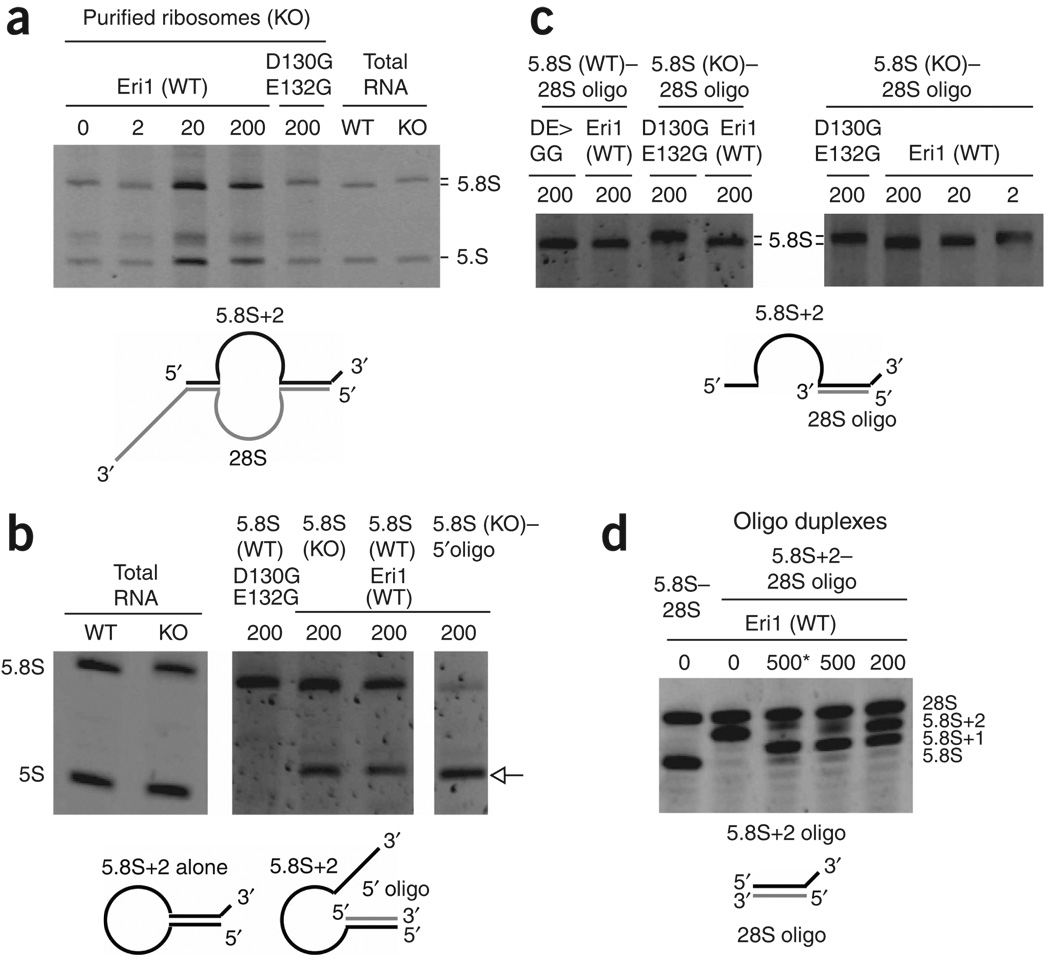
Eri1 is a 3'-to-5' exoribonuclease conserved from fission yeast to humans. Here we show that Eri1 associates with ribosomes and ribosomal RNA (rRNA). Ribosomes from Eri1-deficient mice contain 5.8S rRNA that is aberrantly extended at its 3' end, and Eri1, but not a catalytically inactive mutant, converts this abnormal 5.8S rRNA to the wild-type form in vitro and in cells. In human and murine cells, Eri1 localizes to the cytoplasm and nucleus, with enrichment in the nucleolus, the site of preribosome biogenesis. RNA binding residues in the Eri1 SAP and linker domains promote stable association with rRNA and thereby facilitate 5.8S rRNA 3' end processing. Taken together, our findings indicate that Eri1 catalyzes the final trimming step in 5.8S rRNA processing, functionally and spatially connecting this regulator of RNAi with the basal translation machinery.
Figures
References
-
- Kennedy S, Wang D, Ruvkun G. A conserved siRNA-degrading RNase negatively regulates RNA interference in C. elegans. Nature. 2004;427:645–649. - PubMed
-
- Buhler M, Mohn F, Stalder L, Muhlemann O. Transcriptional silencing of nonsense codon-containing immunoglobulin minigenes. Mol. Cell. 2005;18:307–317. - PubMed
-
- Buhler M, Verdel A, Moazed D. Tethering RITS to a nascent transcript initiates RNAi- and heterochromatin-dependent gene silencing. Cell. 2006;125:873–886. - PubMed
-
- Iida T, Kawaguchi R, Nakayama J. Conserved ribonuclease, Eri1, negatively regulates heterochromatin assembly in fission yeast. Curr. Biol. 2006;16:1459–1464. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
