

Architecture of a serine recombinase-DNA regulatory complex
- PMID: 18439894
- PMCID: PMC2428073
- DOI: 10.1016/j.molcel.2008.02.023
Architecture of a serine recombinase-DNA regulatory complex
Abstract
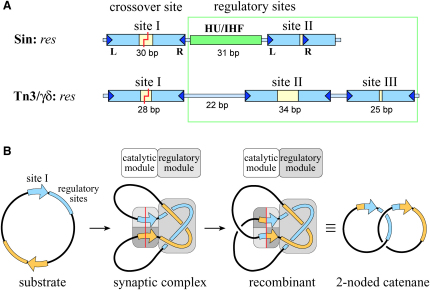
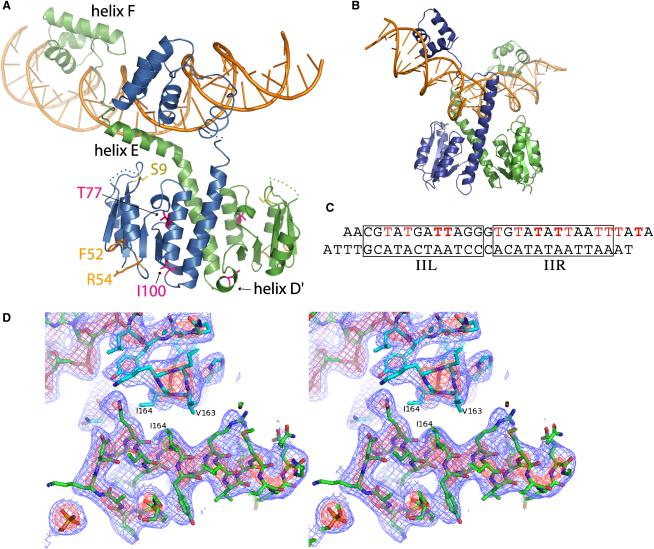
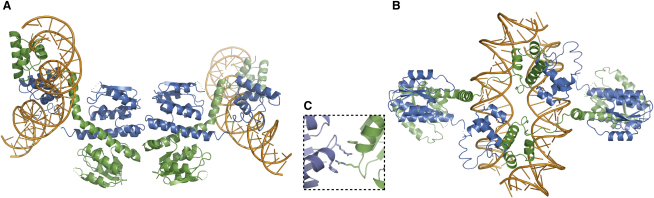
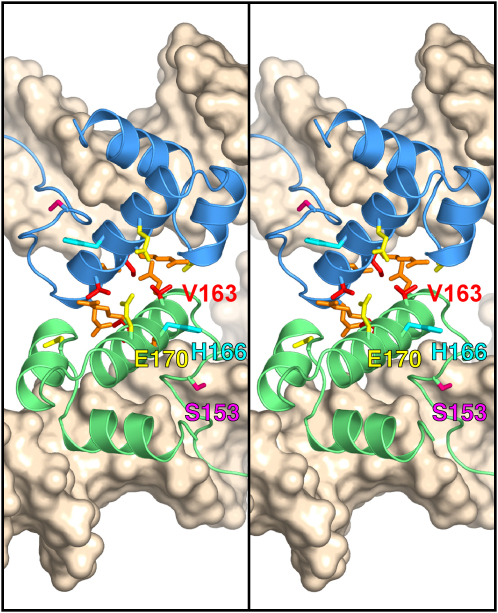
An essential feature of many site-specific recombination systems is their ability to regulate the direction and topology of recombination. Resolvases from the serine recombinase family assemble an interwound synaptic complex that harnesses negative supercoiling to drive the forward reaction and promote recombination between properly oriented sites. To better understand the interplay of catalytic and regulatory functions within these synaptic complexes, we have solved the structure of the regulatory site synapse in the Sin resolvase system. It reveals an unexpected synaptic interface between helix-turn-helix DNA-binding domains that is also highlighted in a screen for synapsis mutants. The tetramer defined by this interface provides the foundation for a robust model of the synaptic complex, assembled entirely from available crystal structures, that gives insight into how the catalytic activity of Sin and other serine recombinases may be regulated.
Figures
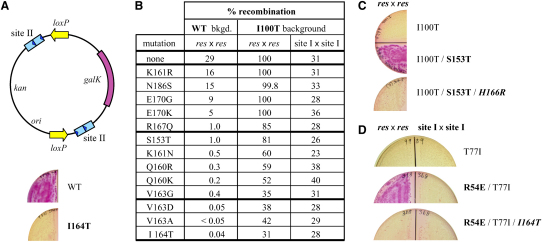
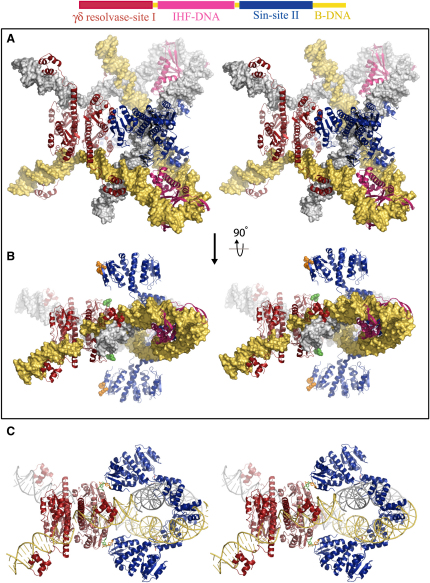
Comment in
-
Assembly of a tightly interwound DNA recombination complex poised for deletion.Structure. 2008 May;16(5):653-5. doi: 10.1016/j.str.2008.04.005. Structure. 2008. PMID: 18462666 No abstract available.
References
-
- Brunger A.T., Adams P.D., Clore G.D., DeLano W.L., Gros P., Grosse-Kunstleve R.W., Jiang J.-S., Kuszewski J., Nilges M., Pannu N.S. Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr. D Biol. Crystallogr. 1998;54:905–921. - PubMed
-
- Calos M.P. The ϕC31 integrase system for gene therapy. Curr. Gene Ther. 2006;6:633–645. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
