

Factor XI homodimer structure is essential for normal proteolytic activation by factor XIIa, thrombin, and factor XIa
- PMID: 18441012
- PMCID: PMC2441546
- DOI: 10.1074/jbc.M802275200
Factor XI homodimer structure is essential for normal proteolytic activation by factor XIIa, thrombin, and factor XIa
Abstract
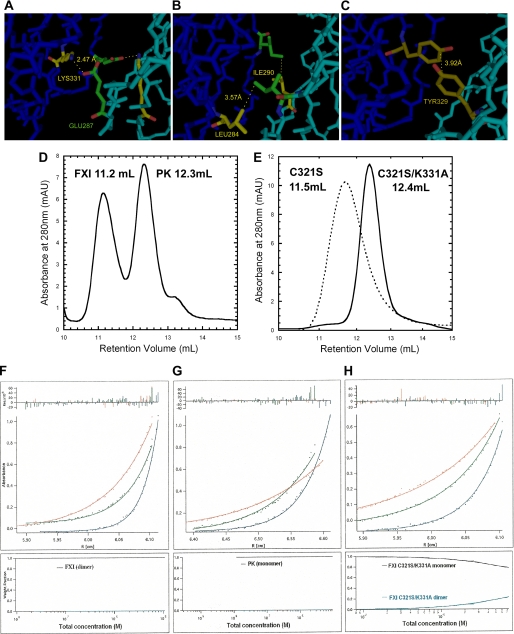
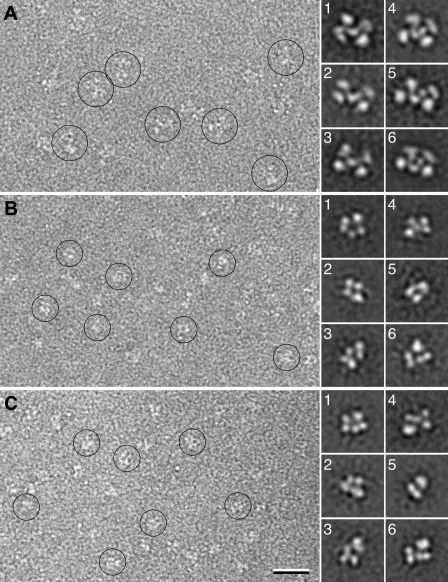
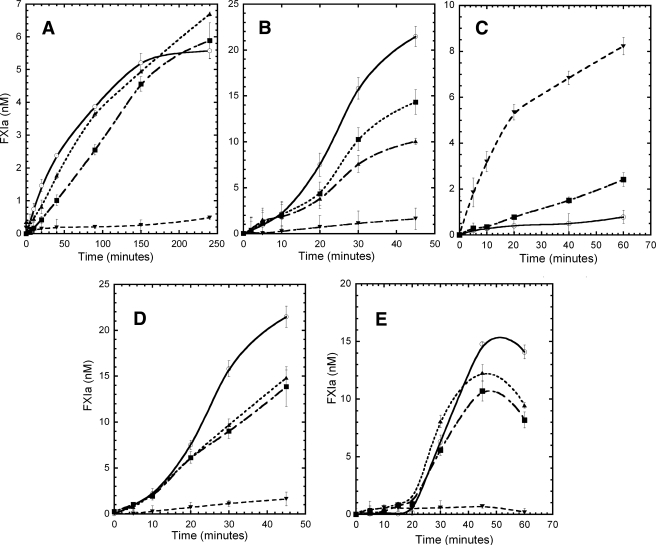
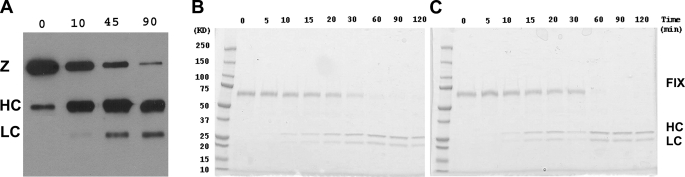
Coagulation factor XI (FXI) is a covalent homodimer consisting of two identical subunits of 80 kDa linked by a disulfide bond formed by Cys-321 within the Apple 4 domain of each subunit. Because FXI(C321S) is a noncovalent dimer, residues within the interface between the two subunits must mediate its homodimeric structure. The crystal structure of FXI demonstrates formation of salt bridges between Lys-331 of one subunit and Glu-287 of the other subunit and hydrophobic interactions at the interface of the Apple 4 domains involving Ile-290, Leu-284, and Tyr-329. FXI(C321S), FXI(C321S,K331A), FXI(C321S,E287A), FXI(C321S,I290A), FXI(C321S,Y329A), FXI(C321S,L284A), FXI(C321S,K331R), and FXI(C321S,H343A) were expressed in HEK293 cells and characterized using size exclusion chromatography, analytical ultracentrifugation, electron microscopy, and functional assays. Whereas FXI(C321S) and FXI(C321S,H343A) existed in monomer/dimer equilibrium (K(d) approximately 40 nm), all other mutants were predominantly monomers with impaired dimer formation by analytical ultracentrifugation (K(d)=3-38 microm). When converted to the active enzyme, FXIa, all the monomeric mutants activated FIX similarly to wild-type dimeric FXIa. In contrast, these monomeric mutants could not be activated efficiently by FXIIa, thrombin, or autoactivation in the presence of dextran sulfate. We conclude that salt bridges formed between Lys-331 of one subunit and Glu-287 of the other together with hydrophobic interactions at the interface, involving residues Ile-290, Leu-284, and Tyr-329, are essential for homodimer formation. The dimeric structure of FXI is essential for normal proteolytic activation of FXI by FXIIa, thrombin, or FXIa either in solution or on an anionic surface but not for FIX activation by FXIa in solution.
Figures
Similar articles
-
The dimeric structure of factor XI and zymogen activation.Blood. 2013 May 9;121(19):3962-9. doi: 10.1182/blood-2012-12-473629. Epub 2013 Mar 20. Blood. 2013. PMID: 23515926 Free PMC article.
-
Dimer dissociation and unfolding mechanism of coagulation factor XI apple 4 domain: spectroscopic and mutational analysis.J Mol Biol. 2007 Mar 23;367(2):558-73. doi: 10.1016/j.jmb.2006.12.066. Epub 2006 Dec 29. J Mol Biol. 2007. PMID: 17257616 Free PMC article.
-
Characterization of a heparin binding site on the heavy chain of factor XI.J Biol Chem. 1998 Nov 20;273(47):31153-9. doi: 10.1074/jbc.273.47.31153. J Biol Chem. 1998. PMID: 9813019
-
The mechanism underlying activation of factor IX by factor XIa.Thromb Res. 2014 May;133 Suppl 1(0 1):S48-51. doi: 10.1016/j.thromres.2014.03.020. Thromb Res. 2014. PMID: 24759143 Free PMC article. Review.
-
Structural and functional features of factor XI.J Thromb Haemost. 2009 Jul;7 Suppl 1(Suppl 1):75-8. doi: 10.1111/j.1538-7836.2009.03414.x. J Thromb Haemost. 2009. PMID: 19630773 Free PMC article. Review.
Cited by
-
The many faces of the contact pathway and their role in thrombosis.J Thromb Thrombolysis. 2011 Jul;32(1):9-20. doi: 10.1007/s11239-011-0578-5. J Thromb Thrombolysis. 2011. PMID: 21404067 Review.
-
Structure and function of factor XI.Blood. 2010 Apr 1;115(13):2569-77. doi: 10.1182/blood-2009-09-199182. Epub 2010 Jan 28. Blood. 2010. PMID: 20110423 Free PMC article. Review.
-
Productive recognition of factor IX by factor XIa exosites requires disulfide linkage between heavy and light chains of factor XIa.J Biol Chem. 2012 Feb 24;287(9):6187-95. doi: 10.1074/jbc.M111.291989. Epub 2011 Dec 29. J Biol Chem. 2012. PMID: 22207756 Free PMC article.
-
Update on the physiology and pathology of factor IX activation by factor XIa.Expert Rev Hematol. 2008 Oct;1(1):87-98. doi: 10.1586/17474086.1.1.87. Expert Rev Hematol. 2008. PMID: 19714257 Free PMC article. Review.
-
An update on factor XI structure and function.Thromb Res. 2018 Jan;161:94-105. doi: 10.1016/j.thromres.2017.10.008. Epub 2017 Oct 10. Thromb Res. 2018. PMID: 29223926 Free PMC article. Review.
References
-
- Naito, K., and Fujikawa, K. (1991) J. Biol. Chem. 266 7353–7358 - PubMed
-
- Gailani, D., and Broze, G. J., Jr. (1991) Science 253 909–912 - PubMed
-
- Fujikawa, K., Legaz, M. E., Kato, H., and Davie, E. W. (1974) Biochemistry 13 4508–4516 - PubMed
-
- Sinha, D., Seaman, F. S., and Walsh, P. N. (1987) Biochemistry 26 3768–3775 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
