

Roles of pneumococcal DivIB in cell division
- PMID: 18441058
- PMCID: PMC2446777
- DOI: 10.1128/JB.00376-08
Roles of pneumococcal DivIB in cell division
Abstract
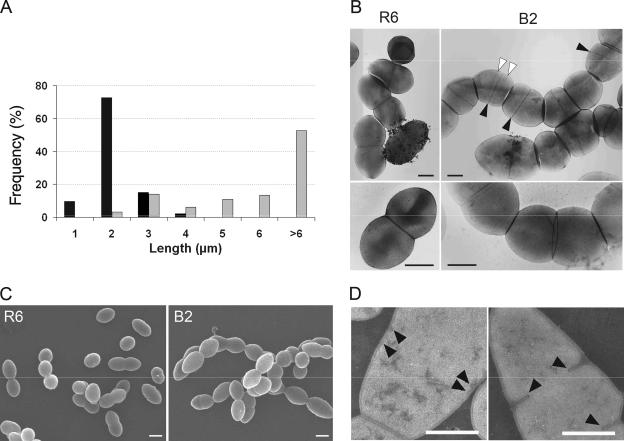
DivIB, also known as FtsQ in gram-negative organisms, is a division protein that is conserved in most eubacteria. DivIB is localized at the division site and forms a complex with two other division proteins, FtsL and DivIC/FtsB. The precise function of these three bitopic membrane proteins, which are central to the division process, remains unknown. We report here the characterization of a divIB deletion mutant of Streptococcus pneumoniae, which is a coccus that divides with parallel planes. Unlike its homologue FtsQ in Escherichia coli, pneumococcal DivIB is not required for growth in rich medium, but the Delta divIB mutant forms chains of diplococci and a small fraction of enlarged cells with defective septa. However, the deletion mutant does not grow in a chemically defined medium. In the absence of DivIB and protein synthesis, the partner FtsL is rapidly degraded, whereas other division proteins are not affected, pointing to a role of DivIB in stabilizing FtsL. This is further supported by the finding that an additional copy of ftsL restores growth of the Delta divIB mutant in defined medium. Functional mapping of the three distinct alpha, beta, and gamma domains of the extracellular region of DivIB revealed that a complete beta domain is required to fully rescue the deletion mutant. DivIB with a truncated beta domain reverts only the chaining phenotype, indicating that DivIB has distinct roles early and late in the division process. Most importantly, the deletion of divIB increases the susceptibility to beta-lactams, more evidently in a resistant strain, suggesting a function in cell wall synthesis.
Figures



Similar articles
-
In vitro reconstitution of a trimeric complex of DivIB, DivIC and FtsL, and their transient co-localization at the division site in Streptococcus pneumoniae.Mol Microbiol. 2005 Jan;55(2):413-24. doi: 10.1111/j.1365-2958.2004.04408.x. Mol Microbiol. 2005. PMID: 15659160
-
Central domain of DivIB caps the C-terminal regions of the FtsL/DivIC coiled-coil rod.J Biol Chem. 2009 Oct 2;284(40):27687-700. doi: 10.1074/jbc.M109.019471. Epub 2009 Jul 27. J Biol Chem. 2009. PMID: 19635793 Free PMC article.
-
Intrinsic instability of the essential cell division protein FtsL of Bacillus subtilis and a role for DivIB protein in FtsL turnover.Mol Microbiol. 2000 Apr;36(2):278-89. doi: 10.1046/j.1365-2958.2000.01857.x. Mol Microbiol. 2000. PMID: 10792716
-
Structural Analysis of the Interaction between the Bacterial Cell Division Proteins FtsQ and FtsB.mBio. 2018 Sep 11;9(5):e01346-18. doi: 10.1128/mBio.01346-18. mBio. 2018. PMID: 30206170 Free PMC article.
-
Evidence from artificial septal targeting and site-directed mutagenesis that residues in the extracytoplasmic β domain of DivIB mediate its interaction with the divisomal transpeptidase PBP 2B.J Bacteriol. 2010 Dec;192(23):6116-25. doi: 10.1128/JB.00783-10. Epub 2010 Sep 24. J Bacteriol. 2010. PMID: 20870765 Free PMC article.
Cited by
-
Regulation of the Peptidoglycan Polymerase Activity of PBP1b by Antagonist Actions of the Core Divisome Proteins FtsBLQ and FtsN.mBio. 2019 Jan 8;10(1):e01912-18. doi: 10.1128/mBio.01912-18. mBio. 2019. PMID: 30622193 Free PMC article.
-
Synergism between Corynebacterium and Streptococcus sanguinis reveals new interactions between oral commensals.ISME J. 2020 May;14(5):1154-1169. doi: 10.1038/s41396-020-0598-2. Epub 2020 Feb 4. ISME J. 2020. PMID: 32020052 Free PMC article.
-
Multiple interaction domains in FtsL, a protein component of the widely conserved bacterial FtsLBQ cell division complex.J Bacteriol. 2010 Jun;192(11):2757-68. doi: 10.1128/JB.01609-09. Epub 2010 Apr 2. J Bacteriol. 2010. PMID: 20363951 Free PMC article.
-
Bacteriocin protein BacL1 of Enterococcus faecalis targets cell division loci and specifically recognizes L-Ala2-cross-bridged peptidoglycan.J Bacteriol. 2015 Jan;197(2):286-95. doi: 10.1128/JB.02203-14. Epub 2014 Nov 3. J Bacteriol. 2015. PMID: 25368300 Free PMC article.
-
The Cell Wall of Streptococcus pneumoniae.Microbiol Spectr. 2019 May;7(3):10.1128/microbiolspec.gpp3-0018-2018. doi: 10.1128/microbiolspec.GPP3-0018-2018. Microbiol Spectr. 2019. PMID: 31172911 Free PMC article. Review.
References
-
- Angert, E. R. 2005. Alternatives to binary fission in bacteria. Nat. Rev. Microbiol. 3214-224. - PubMed
-
- Bramkamp, M., L. Weston, R. A. Daniel, and J. Errington. 2006. Regulated intramembrane proteolysis of FtsL protein and the control of cell division in Bacillus subtilis. Mol. Microbiol. 62580-591. - PubMed
-
- Briegel, A., D. P. Dias, Z. Li, R. B. Jensen, A. S. Frangakis, and G. J. Jensen. 2006. Multiple large filament bundles observed in Caulobacter crescentus by electron cryotomography. Mol. Microbiol. 625-14. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
