

Chromosomal toxin-antitoxin systems may act as antiaddiction modules
- PMID: 18441063
- PMCID: PMC2446810
- DOI: 10.1128/JB.00357-08
Chromosomal toxin-antitoxin systems may act as antiaddiction modules
Abstract
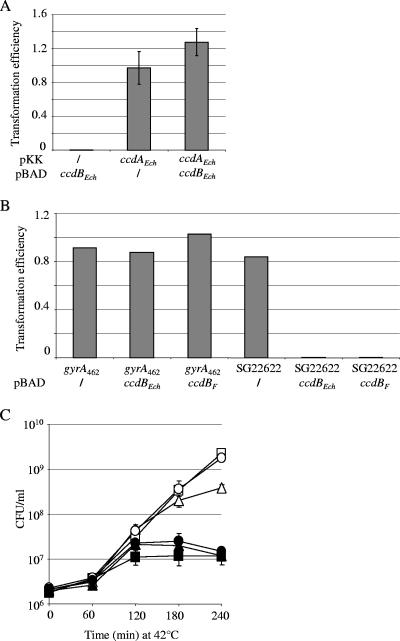
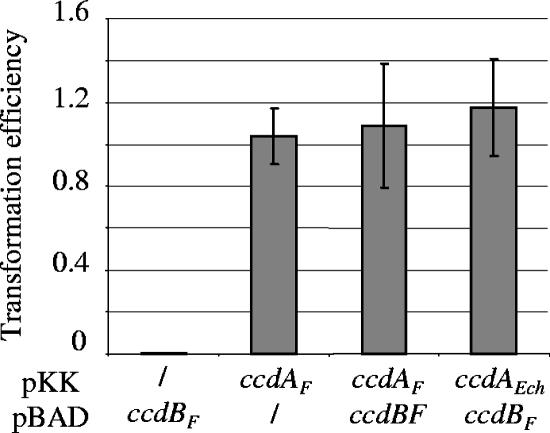
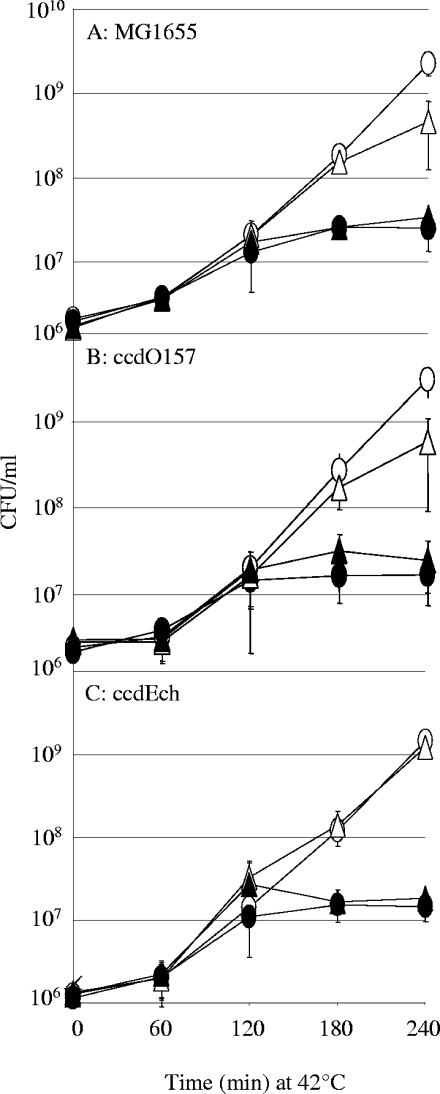
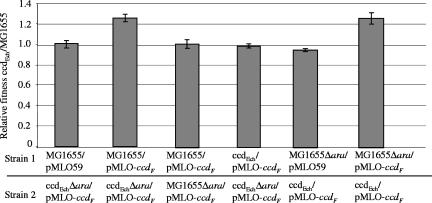
Toxin-antitoxin (TA) systems are widespread among bacterial chromosomes and mobile genetic elements. Although in plasmids TA systems have a clear role in their vertical inheritance by selectively killing plasmid-free daughter cells (postsegregational killing or addiction phenomenon), the physiological role of chromosomally encoded ones remains under debate. The assumption that chromosomally encoded TA systems are part of stress response networks and/or programmed cell death machinery has been called into question recently by the observation that none of the five canonical chromosomally encoded TA systems in the Escherichia coli chromosome seem to confer any selective advantage under stressful conditions (V. Tsilibaris, G. Maenhaut-Michel, N. Mine, and L. Van Melderen, J. Bacteriol. 189:6101-6108, 2007). Their prevalence in bacterial chromosomes indicates that they might have been acquired through horizontal gene transfer. Once integrated in chromosomes, they might in turn interfere with their homologues encoded by mobile genetic elements. In this work, we show that the chromosomally encoded Erwinia chrysanthemi ccd (control of cell death) (ccd(Ech)) system indeed protects the cell against postsegregational killing mediated by its F-plasmid ccd (ccd(F)) homologue. Moreover, competition experiments have shown that this system confers a fitness advantage under postsegregational conditions mediated by the ccd(F) system. We propose that ccd(Ech) acts as an antiaddiction module and, more generally, that the integration of TA systems in bacterial chromosomes could drive the evolution of plasmid-encoded ones and select toxins that are no longer recognized by the antiaddiction module.
Figures
Similar articles
-
Functional interactions between coexisting toxin-antitoxin systems of the ccd family in Escherichia coli O157:H7.J Bacteriol. 2007 Apr;189(7):2712-9. doi: 10.1128/JB.01679-06. Epub 2007 Jan 26. J Bacteriol. 2007. PMID: 17259320 Free PMC article.
-
Contribution of the Chromosomal ccdAB Operon to Bacterial Drug Tolerance.J Bacteriol. 2017 Sep 5;199(19):e00397-17. doi: 10.1128/JB.00397-17. Print 2017 Oct 1. J Bacteriol. 2017. PMID: 28674066 Free PMC article.
-
Toxin-antitoxin systems in the mobile genome of Acidithiobacillus ferrooxidans.PLoS One. 2014 Nov 10;9(11):e112226. doi: 10.1371/journal.pone.0112226. eCollection 2014. PLoS One. 2014. PMID: 25384039 Free PMC article.
-
Type II Toxin-Antitoxin Systems: Evolution and Revolutions.J Bacteriol. 2020 Mar 11;202(7):e00763-19. doi: 10.1128/JB.00763-19. Print 2020 Mar 11. J Bacteriol. 2020. PMID: 31932311 Free PMC article. Review.
-
Bacterial toxin-antitoxin systems: more than selfish entities?PLoS Genet. 2009 Mar;5(3):e1000437. doi: 10.1371/journal.pgen.1000437. Epub 2009 Mar 27. PLoS Genet. 2009. PMID: 19325885 Free PMC article. Review.
Cited by
-
Ciprofloxacin causes persister formation by inducing the TisB toxin in Escherichia coli.PLoS Biol. 2010 Feb 23;8(2):e1000317. doi: 10.1371/journal.pbio.1000317. PLoS Biol. 2010. PMID: 20186264 Free PMC article.
-
A toxin-antidote selfish element increases fitness of its host.Elife. 2023 Oct 24;12:e81640. doi: 10.7554/eLife.81640. Elife. 2023. PMID: 37874324 Free PMC article.
-
Insights into Transcriptional Repression of the Homologous Toxin-Antitoxin Cassettes yefM-yoeB and axe-txe.Int J Mol Sci. 2020 Nov 28;21(23):9062. doi: 10.3390/ijms21239062. Int J Mol Sci. 2020. PMID: 33260607 Free PMC article.
-
Assessing the Role of Bacterial Innate and Adaptive Immunity as Barriers to Conjugative Plasmids.Mol Biol Evol. 2024 Oct 4;41(10):msae207. doi: 10.1093/molbev/msae207. Mol Biol Evol. 2024. PMID: 39382385 Free PMC article.
-
To be or not to be: regulation of restriction-modification systems and other toxin-antitoxin systems.Nucleic Acids Res. 2014 Jan;42(1):70-86. doi: 10.1093/nar/gkt711. Epub 2013 Aug 13. Nucleic Acids Res. 2014. PMID: 23945938 Free PMC article. Review.
References
-
- Barrangou, R., C. Fremaux, H. Deveau, M. Richards, P. Boyaval, S. Moineau, D. A. Romero, and P. Horvath. 2007. CRISPR provides acquired resistance against viruses in prokaryotes. Science 3151709-1712. - PubMed
-
- Bernard, P., and M. Couturier. 1992. Cell killing by the F plasmid CcdB protein involves poisoning of DNA-topoisomerase II complexes. J. Mol. Biol. 226735-745. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
