

Entry to "formula tunnel" revealed by SLC4A4 human mutation and structural model
- PMID: 18441326
- PMCID: PMC2440602
- DOI: 10.1074/jbc.M709819200
Entry to "formula tunnel" revealed by SLC4A4 human mutation and structural model
Abstract
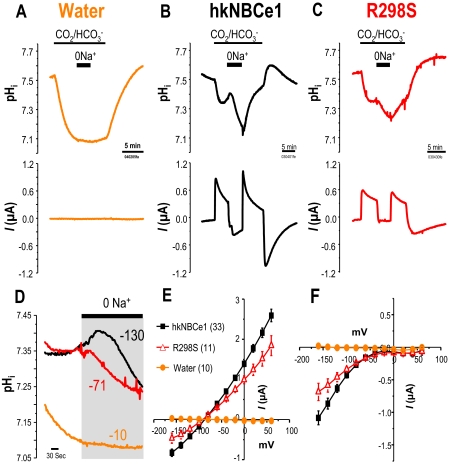
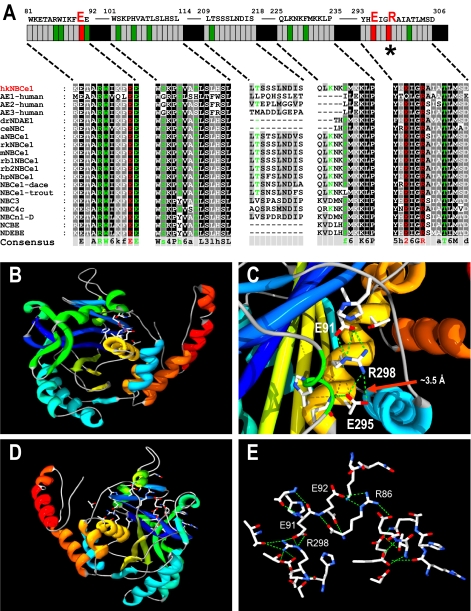
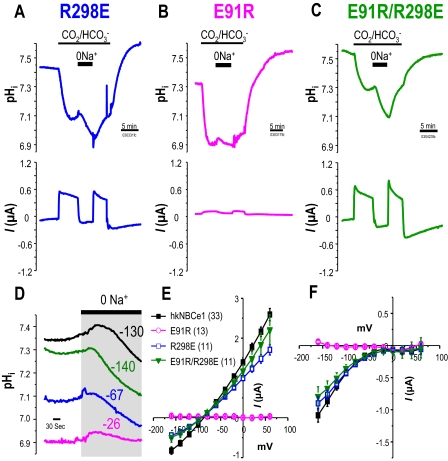
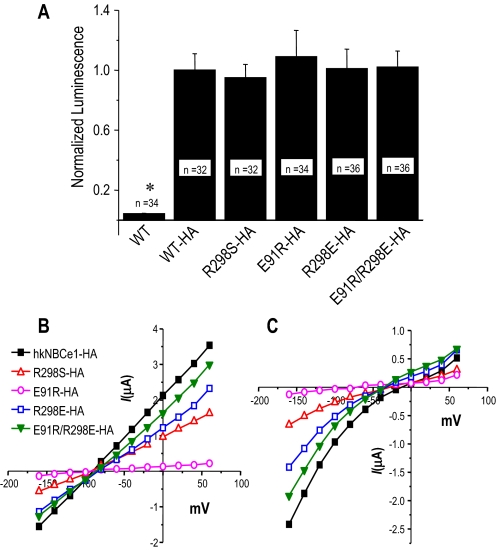
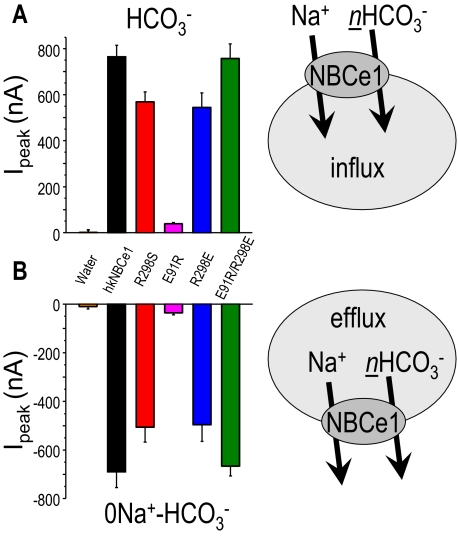
Glaucoma, cataracts, and proximal renal tubular acidosis are diseases caused by point mutations in the human electrogenic Na(+) bicarbonate cotransporter (NBCe1/SLC4A4) (1, 2). One such mutation, R298S, is located in the cytoplasmic N-terminal domain of NBCe1 and has only moderate (75%) function. As SLC transporters have high similarity in their membrane and N-terminal primary sequences, we homology-modeled NBCe1 onto the crystal structure coordinates of Band 3(AE1) (3). Arg-298 is predicted to be located in a solvent-inaccessible subsurface pocket and to associate with Glu-91 or Glu-295 via H-bonding and charge-charge interactions. We perturbed these putative interactions between Glu-91 and Arg-298 by site-directed mutagenesis and used expression in Xenopus oocyte to test our structural model. Mutagenesis of either residue resulted in reduced transport function. Function was "repaired" by charge reversal (E91R/R298E), implying that these two residues are interchangeable and interdependent. These results contrast the current understanding of the AE1 N terminus as protein-binding sites and propose that hkNBCe1 (and other SLC4) cytoplasmic N termini play roles in controlling HCO(3)(-) permeation.
Figures
Similar articles
-
A substrate access tunnel in the cytosolic domain is not an essential feature of the solute carrier 4 (SLC4) family of bicarbonate transporters.J Biol Chem. 2013 Nov 22;288(47):33848-33860. doi: 10.1074/jbc.M113.511865. Epub 2013 Oct 11. J Biol Chem. 2013. PMID: 24121512 Free PMC article.
-
A novel missense mutation in the sodium bicarbonate cotransporter (NBCe1/SLC4A4) causes proximal tubular acidosis and glaucoma through ion transport defects.J Biol Chem. 2004 Dec 10;279(50):52238-46. doi: 10.1074/jbc.M406591200. Epub 2004 Oct 7. J Biol Chem. 2004. PMID: 15471865
-
Proximal renal tubular acidosis and ocular pathology: a novel missense mutation in the gene (SLC4A4) for sodium bicarbonate cotransporter protein (NBCe1).Mol Vis. 2006 Apr 10;12:324-30. Mol Vis. 2006. PMID: 16636648
-
[Physiology and pathophysiology of Na⁺/HCO₃⁻ cotransporter NBCe1].Sheng Li Xue Bao. 2012 Dec 25;64(6):729-40. Sheng Li Xue Bao. 2012. PMID: 23258339 Review. Chinese.
-
NBCe1 as a model carrier for understanding the structure-function properties of Na⁺ -coupled SLC4 transporters in health and disease.Pflugers Arch. 2014 Aug;466(8):1501-16. doi: 10.1007/s00424-014-1448-8. Epub 2014 Feb 11. Pflugers Arch. 2014. PMID: 24515290 Free PMC article. Review.
Cited by
-
Structure, function, and regulation of the SLC4 NBCe1 transporter and its role in causing proximal renal tubular acidosis.Curr Opin Nephrol Hypertens. 2013 Sep;22(5):572-83. doi: 10.1097/MNH.0b013e328363ff43. Curr Opin Nephrol Hypertens. 2013. PMID: 23917030 Free PMC article. Review.
-
NBCe1A dimer assemble visualized by bimolecular fluorescence complementation.Am J Physiol Renal Physiol. 2014 Mar 15;306(6):F672-80. doi: 10.1152/ajprenal.00284.2013. Epub 2014 Jan 29. Am J Physiol Renal Physiol. 2014. PMID: 24477681 Free PMC article.
-
A substrate access tunnel in the cytosolic domain is not an essential feature of the solute carrier 4 (SLC4) family of bicarbonate transporters.J Biol Chem. 2013 Nov 22;288(47):33848-33860. doi: 10.1074/jbc.M113.511865. Epub 2013 Oct 11. J Biol Chem. 2013. PMID: 24121512 Free PMC article.
-
The colorful mantle of the giant clam Tridacna squamosa expresses a homolog of electrogenic sodium: Bicarbonate cotransporter 2 that mediates the supply of inorganic carbon to photosynthesizing symbionts.PLoS One. 2021 Oct 15;16(10):e0258519. doi: 10.1371/journal.pone.0258519. eCollection 2021. PLoS One. 2021. PMID: 34653199 Free PMC article.
-
Proximal renal tubular acidosis mediated by mutations in NBCe1-A: unraveling the transporter's structure-functional properties.Front Physiol. 2013 Dec 19;4:350. doi: 10.3389/fphys.2013.00350. Front Physiol. 2013. PMID: 24391589 Free PMC article. Review.
References
-
- Igarashi, T., Inatomi, J., Sekine, T., Cha, S. H., Kanai, Y., Kunimi, M., Tsukamoto, K., Satoh, H., Shimadzu, M., Tozawa, F., Mori, T., Shiobara, M., Seki, G., and Endou, H. (1999) Nat. Genet. 23 264-266 - PubMed
-
- Romero, M. F., and Smith, B. L. (2002) FASEB J. 16 a52-a53
-
- Zhang, D., Kiyatkin, A., Bolin, J. T., and Low, P. S. (2000) Blood 96 2925-2933 - PubMed
-
- Alper, S. L. (2002) Annu. Rev. Physiol. 64 899-923 - PubMed
-
- Romero, M. F. (2005) Curr. Opin. Nephrol. Hypertens. 14 495-501 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
