

Investigation of the role of breast cancer resistance protein (Bcrp/Abcg2) on pharmacokinetics and central nervous system penetration of abacavir and zidovudine in the mouse
- PMID: 18443033
- PMCID: PMC5091086
- DOI: 10.1124/dmd.108.020974
Investigation of the role of breast cancer resistance protein (Bcrp/Abcg2) on pharmacokinetics and central nervous system penetration of abacavir and zidovudine in the mouse
Abstract
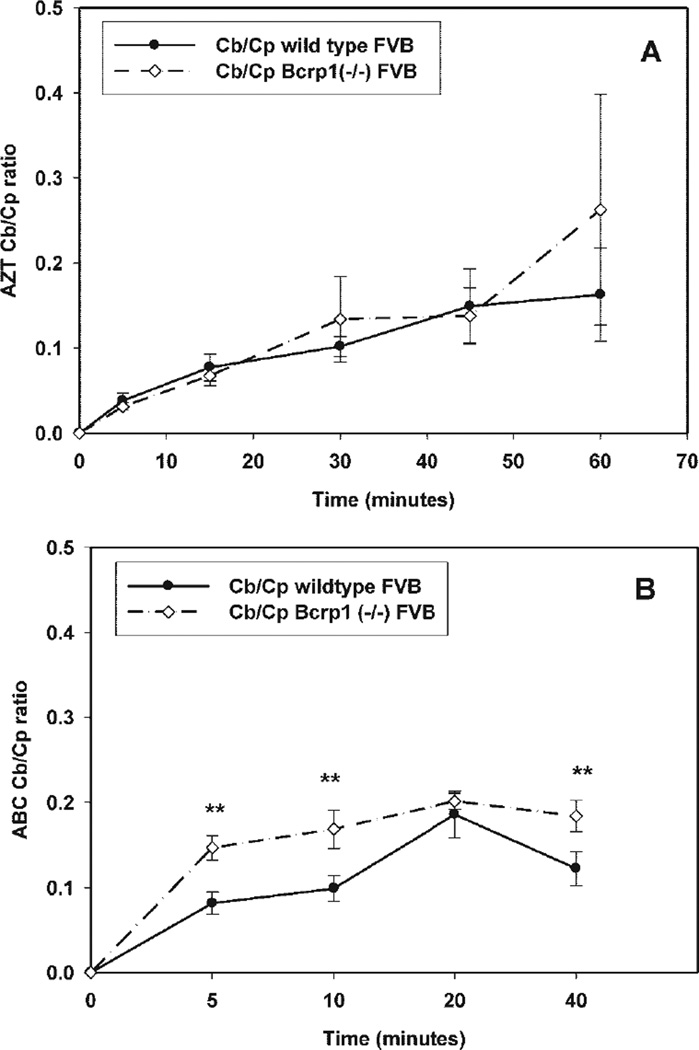
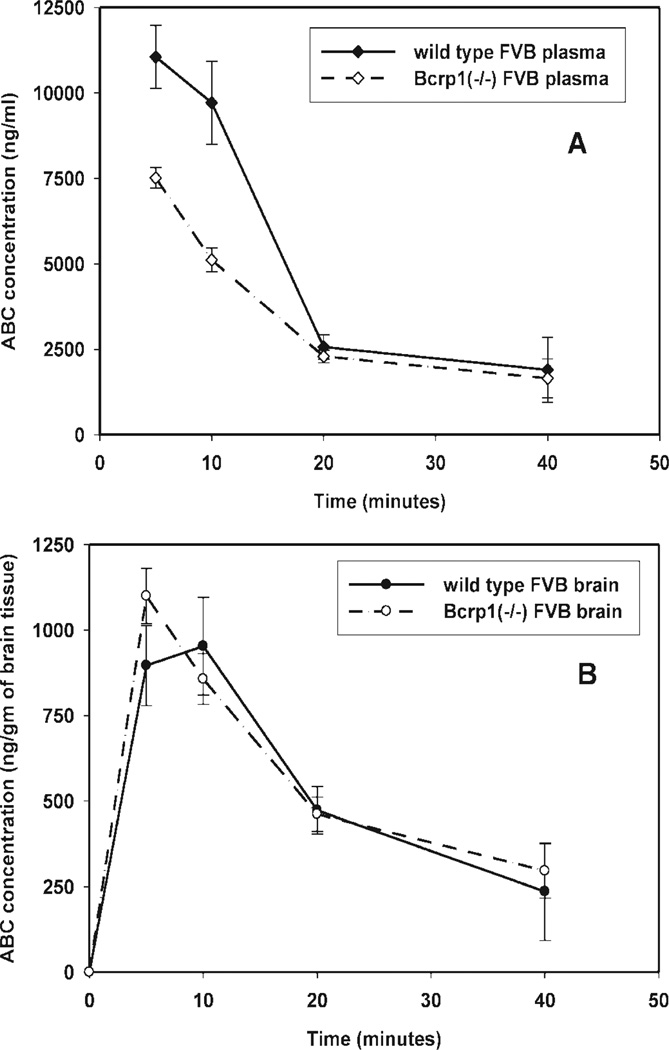
Many anti-human immunodeficiency virus 1 nucleoside reverse-transcriptase inhibitors have low central nervous system (CNS) distribution due in part to active efflux transport at the blood-brain barrier. We have previously shown that zidovudine (AZT) and abacavir (ABC) are in vitro substrates for the efflux transport protein breast cancer resistance protein (Bcrp) 1. We evaluated the influence of Bcrp1 on plasma pharmacokinetics and brain penetration of zidovudine and abacavir in wild-type and Bcrp1-deficient (Bcrp1-/-) FVB mice. There was no difference in either area under the concentration-time profiles for plasma (AUC(plasma)) or brain (AUC(brain)) for zidovudine between the wild-type and Bcrp1-/- mice. The AUC(plasma) of abacavir was 20% lower in the Bcrp1-/- mice, whereas the AUC(brain) was 20% greater. This difference resulted in a 1.5-fold increase in abacavir brain exposure in the Bcrp1-/- mice. The effect of selective and nonselective transport inhibitors on the ABC brain/plasma ratio at a single time point was evaluated. 3-(6-Isobutyl-9-methoxy-1,4-dioxo-1,2,3,4,6, 7,12,12a-octahydropyrazino[1',2':1,6]pyrido[3,4-b]indol-3-yl)-propionicacid tert-butyl ester (Ko143), N[4[2-(6,7-dimethoxy-3,4-dihydro-1H-isoquinolin-2-yl)ethyl]phenyl]-5-methoxy-9-oxo-10H-acridine-4-carboxamide (GF120918), probenecid, and Pluronic P85 increased abacavir plasma concentrations in the wild-type mice. Abacavir plasma concentrations in Bcrp1-/- mice were increased by (R)-4-((1aR,6R,10bS)-1,2-difluoro-1,1a,6,10b-tetrahydrodibenzo (a,e)cyclopropa(c)cycloheptan-6-yl)-alpha-((5-quinoloyloxy)methyl)-1-piperazineethanol trihydrochloride (LY335979), GF120918, and probenecid, but not by Ko143. Brain/plasma concentration ratios in both the wild-type and Bcrp1-/- mice were increased by the P-glycoprotein inhibitors LY335979 and GF120918, but not by BCRP-selective inhibitors. These data indicate that deletion of Bcrp1 has little influence on the pharmacokinetics or brain penetration of AZT. However, for abacavir, deletion of Bcrp1 reduces plasma exposure and enhances brain penetration. These findings suggest that Bcrp1 does not play a significant role in limiting the CNS distribution of zidovudine and abacavir; however, brain penetration of abacavir is dependent on P-glycoprotein-mediated efflux.
Figures
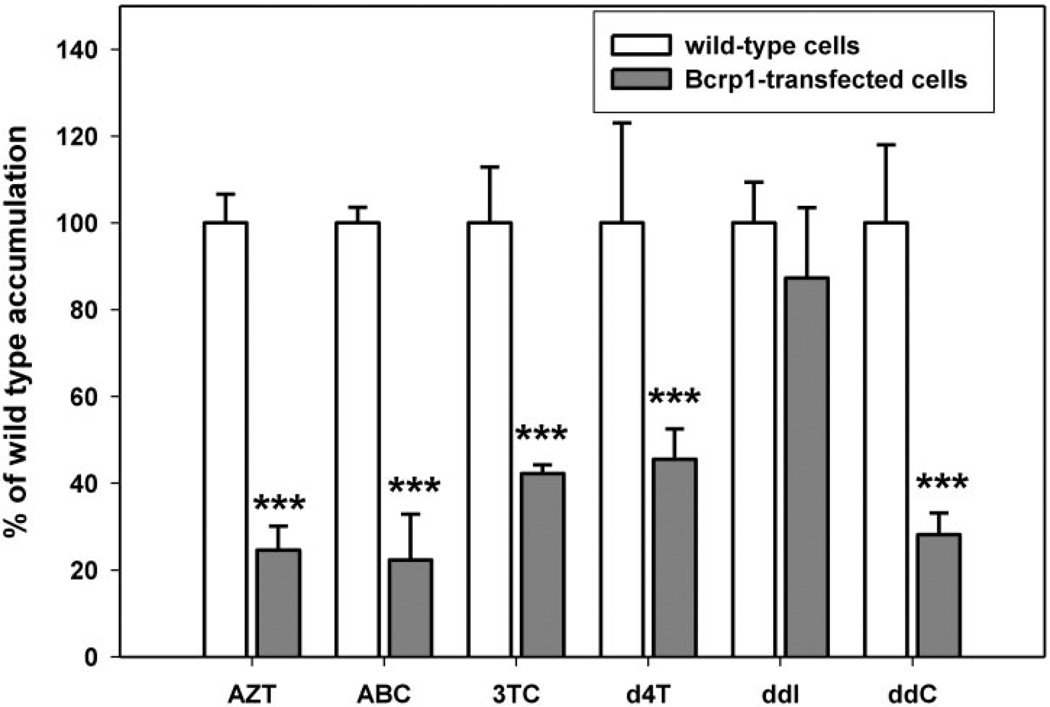
 ) MDCKII cells. Results are expressed as means ± S.D.; n = 3. ***, p < 0.001, compared with the wild-type control group.
) MDCKII cells. Results are expressed as means ± S.D.; n = 3. ***, p < 0.001, compared with the wild-type control group.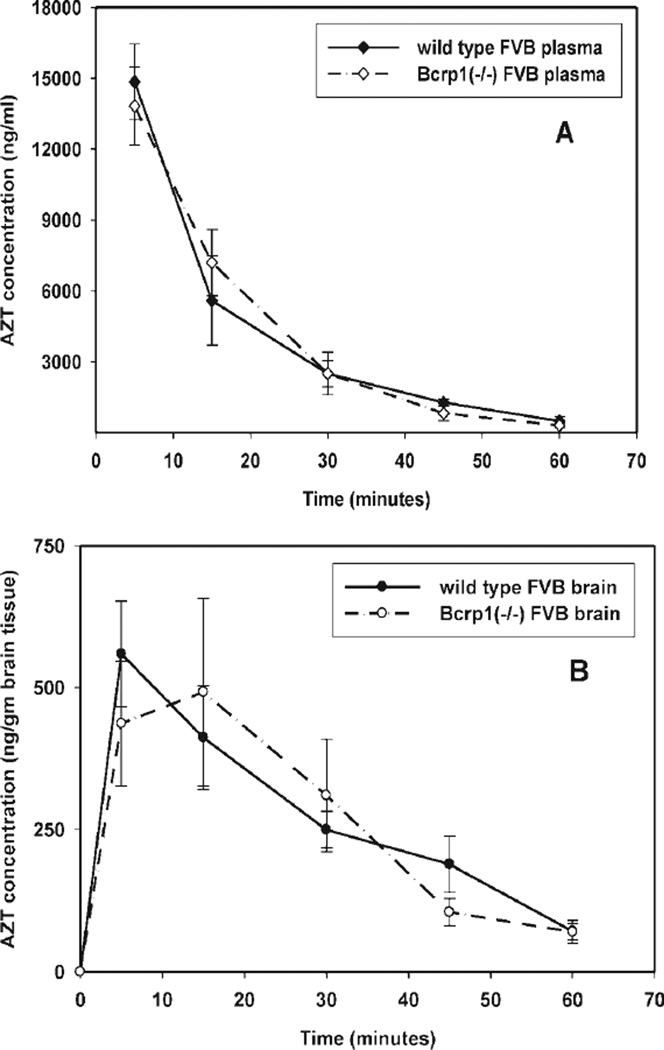
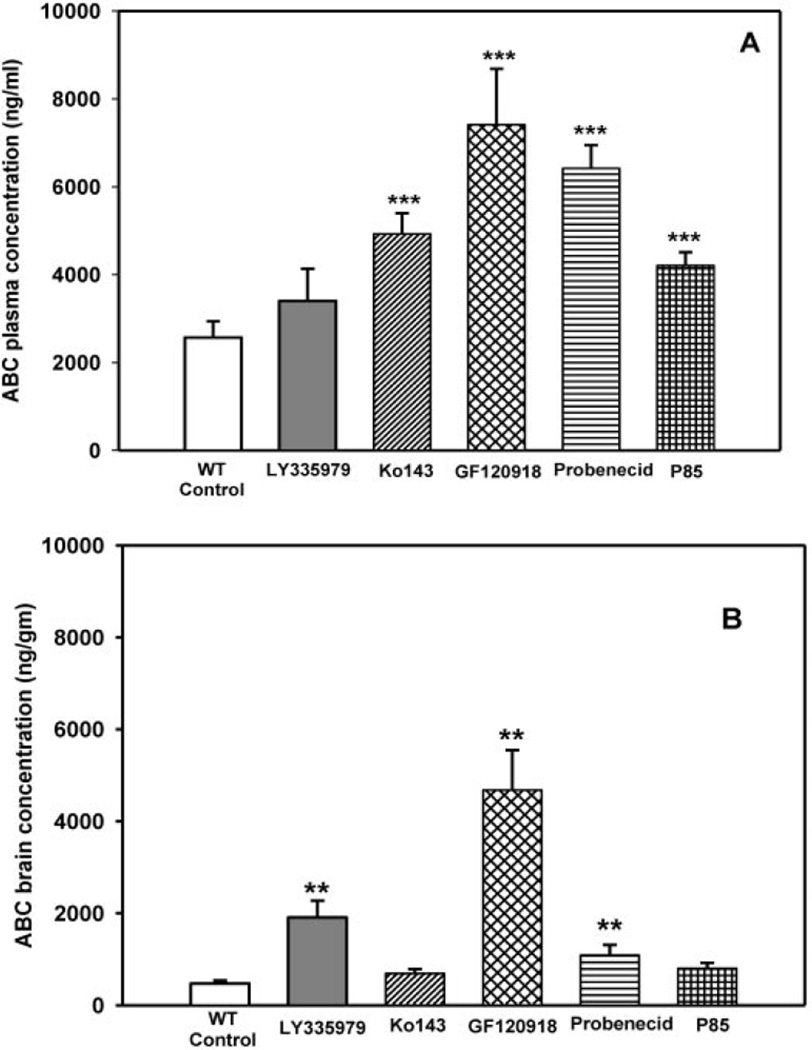 , 25 mg/kg LY335979; ▨, 1 mg/kg Ko413; is ▩, 10 mg/kg GF120918; ▤, 200 mg/kg probenecid; ▦, 20 mg/kg Pluronic P85. Results are expressed as means ± S.D.; n = 4. ***, p < 0.001; **, p < 0.01 compared with the respective wild-type controls.
, 25 mg/kg LY335979; ▨, 1 mg/kg Ko413; is ▩, 10 mg/kg GF120918; ▤, 200 mg/kg probenecid; ▦, 20 mg/kg Pluronic P85. Results are expressed as means ± S.D.; n = 4. ***, p < 0.001; **, p < 0.01 compared with the respective wild-type controls.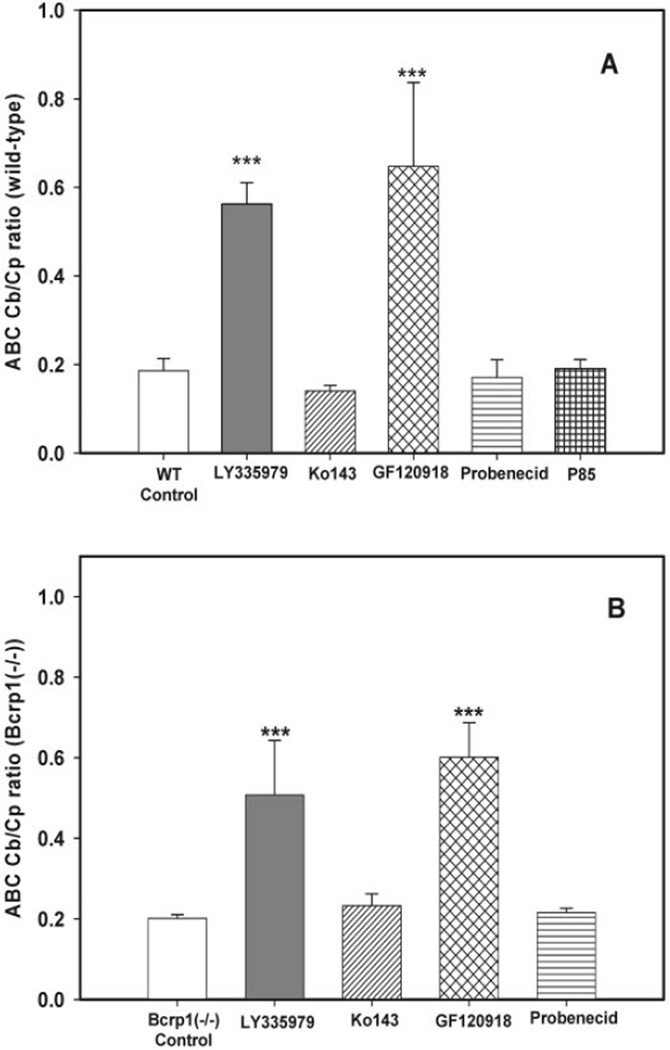 , 25 mg/kg LY335979; ▨, 1 mg/kg Ko413; ▩, 10 mg/kg GF120918; ▤, 200 mg/kg probenecid; ▦, 20 mg/kg Pluronic P85. Results are expressed as means ± S.D.; n = 4. ***, p < 0.001. compared with the respective wild-type and Bcrp1−/− controls.
, 25 mg/kg LY335979; ▨, 1 mg/kg Ko413; ▩, 10 mg/kg GF120918; ▤, 200 mg/kg probenecid; ▦, 20 mg/kg Pluronic P85. Results are expressed as means ± S.D.; n = 4. ***, p < 0.001. compared with the respective wild-type and Bcrp1−/− controls.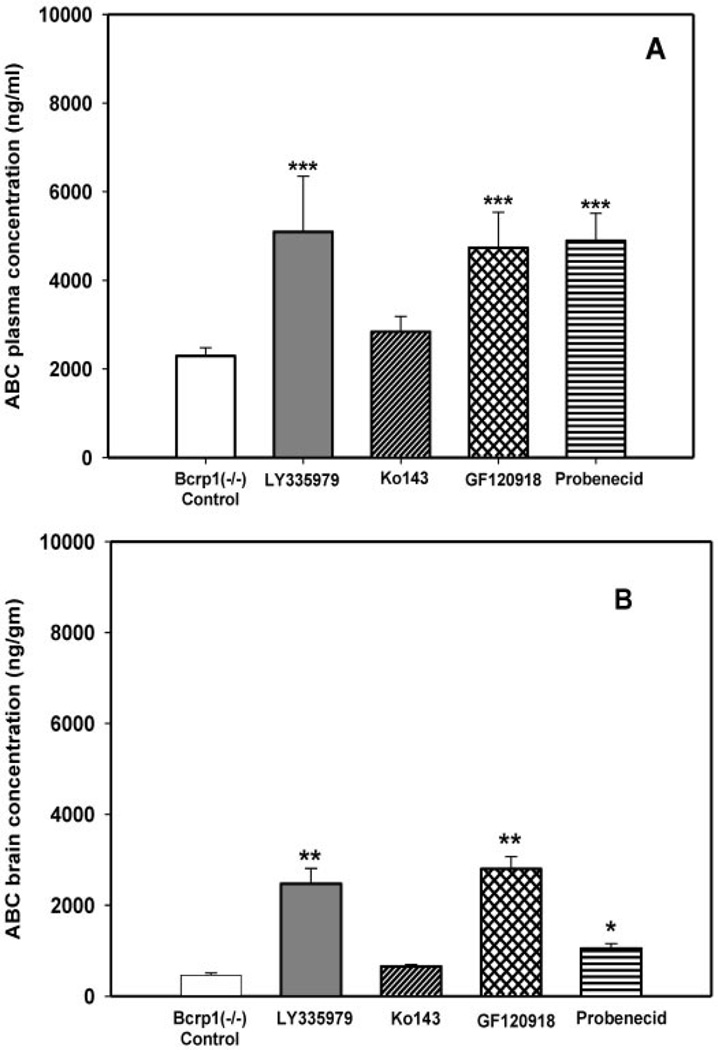 , 25 mg/kg LY335979; ▨, 1 mg/kg Ko413; ▩, 10 mg/kg GF120918; ▤, 200 mg/kg probenecid; ▦, 20 mg/kg Pluronic P85. Results are expressed as means ± S.D.; n = 4. ***, p < 0.001; **, p < 0.01 compared with the respective Bcrp1−/− controls.
, 25 mg/kg LY335979; ▨, 1 mg/kg Ko413; ▩, 10 mg/kg GF120918; ▤, 200 mg/kg probenecid; ▦, 20 mg/kg Pluronic P85. Results are expressed as means ± S.D.; n = 4. ***, p < 0.001; **, p < 0.01 compared with the respective Bcrp1−/− controls.References
-
- Bailer AJ. Testing for the equality of area under the curves when using destructive measurement techniques. J Pharmacokinet Biopharm. 1988;16:303–309. - PubMed
-
- Batrakova EV, Miller DW, Li S, Alakhov VY, Kabanov AV, Elmquist WF. Pluronic P85 enhances the delivery of digoxin to the brain: in vitro and in vivo studies. J Pharmacol Exp Ther. 2001;296:551–557. - PubMed
-
- Bihorel S, Camenisch G, Lemaire M, Scherrmann JM. Modulation of the brain distribution of imatinib and its metabolites in mice by valspodar, zosuquidar and elacridar. Pharm Res. 2007;24:1720–1728. - PubMed
-
- Cooray HC, Blackmore CG, Maskell L, Barrand MA. Localisation of breast cancer resistance protein in microvessel endothelium of human brain. Neuroreport. 2002;13:2059–2063. - PubMed
-
- Cordon-Cardo C, O’Brien JP, Boccia J, Casals D, Bertino JR, Melamed MR. Expression of the multidrug resistance gene product (P-glycoprotein) in human normal and tumor tissues. J Histochem Cytochem. 1990;38:1277–1287. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
