

REIC/Dkk-3 induces cell death in human malignant glioma
- PMID: 18443132
- PMCID: PMC2563047
- DOI: 10.1215/15228517-2008-016
REIC/Dkk-3 induces cell death in human malignant glioma
Abstract
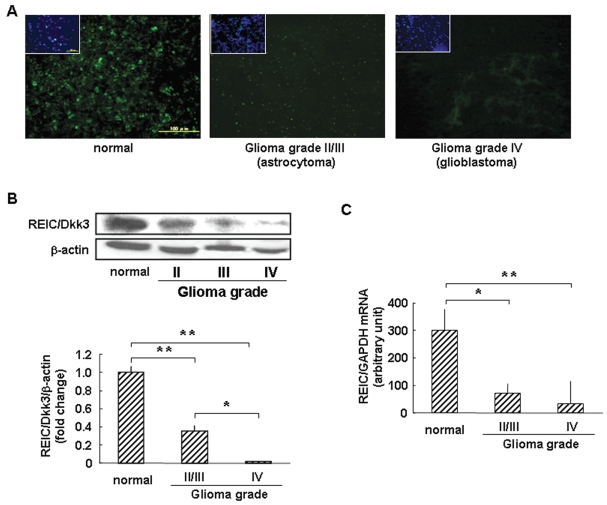
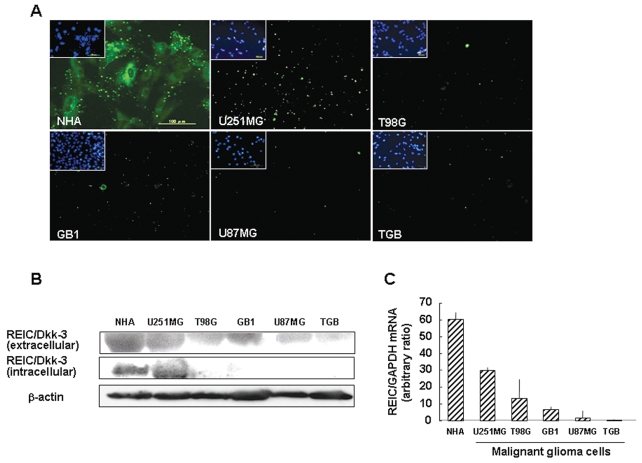
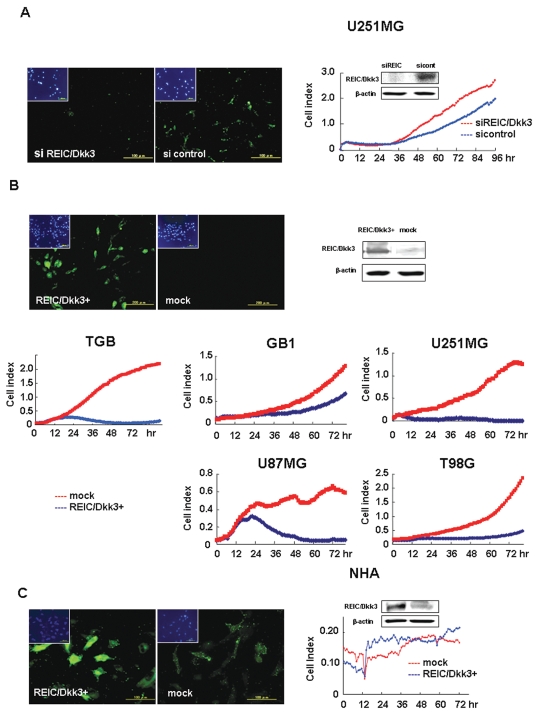
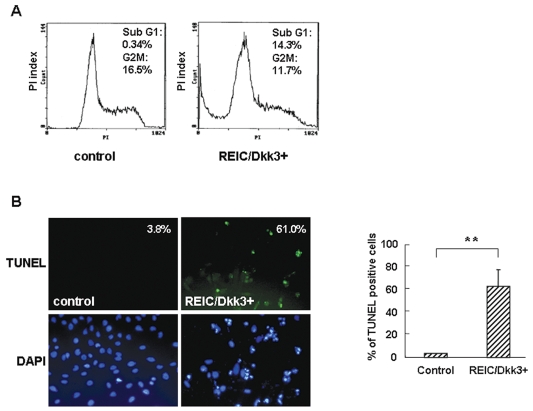
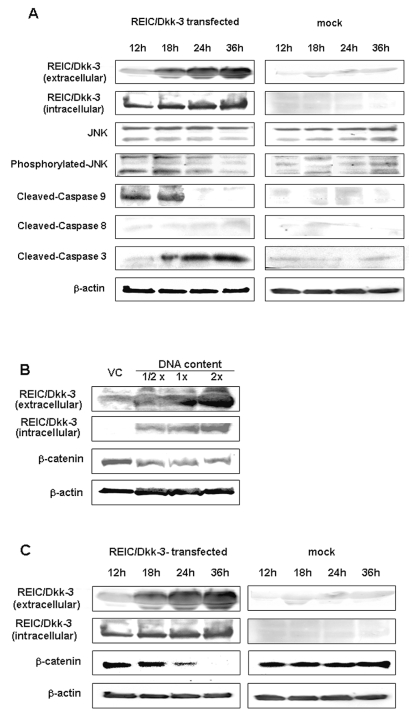
The progression of glioma to more malignant phenotypes results from the stepwise accumulation of genetic alterations and the consequent disruption of the apoptotic pathway and augmentation of survival signaling. REIC/Dkk-3, a member of the human Dickkopf (Dkk) family, plays a role as a suppressor of the growth of several human cancers; however, to date it has not been identified in brain tumors. We compared the gene and protein expression of REIC/Dkk-3 in human malignant glioma and normal brain tissues using quantitative real-time PCR, Western blotting, and immunohistochemistry. We also performed small interfering REIC/Dkk-3 (siREIC/Dkk-3) knockdown and REIC/Dkk-3 overexpression experiments to examine the role of REIC/Dkk-3 in human malignant glioma cells in vitro. In brain tissue from patients with malignant glioma, the gene and protein expression of REIC/Dkk-3 was lower than in normal brain tissue and was related to the malignancy grade. In the primary glioblastoma cell line, REIC/Dkk-3 transfection led to apoptosis owing to the activation of phosphorylated JUN, caspase-9, and caspase-3 and the reduction of beta-catenin; in REIC/Dkk-3 knockdown experiments, cell growth was augmented. Our results suggest that REIC/Dkk-3 regulates the growth and survival of these cells in a caspase-dependent and -independent way via modification of the Wnt signaling pathway. Our work is the first documentation that the gene and protein expression of REIC/Dkk-3 is down-regulated in human malignant glioma. Our demonstration of the mechanisms underlying REIC/Dkk-3-induced cell death indicates that REIC/Dkk-3 plays a pivotal role in the biology of human malignant glioma and suggests that REIC/Dkk-3 is a promising candidate for molecular target therapy.
Figures
References
-
- Farhadi MR, Capelle HH, Erber R, Ullrich A, Vajkoczy P. Combined inhibition of vascular endothelial growth factor and platelet-derived growth factor signaling: effects on the angiogenesis, microcirculation, and growth of orthotopic malignant gliomas. J Neurosurg. 2005;102:363 – 370. - PubMed
-
- Stewart LA. Chemotherapy in adult high-grade glioma: a systematic review and meta-analysis of individual patient data from 12 randomised trials. Lancet. 2002;359:1011 – 1018. - PubMed
-
- Committee of Brain Tumor Registry of Japan. Report of Brain Tumor Registry of Japan (1969 – 1996) Neurol Med Chir (Tokyo) 2003;43 (suppl i – vii):1 – 111. - PubMed
-
- Cavenee WK. Accumulation of genetic defects during astrocytoma progression. Cancer. 1992;70:1788 – 1793. - PubMed
-
- Wang J, Shou J, Chen X. Dickkopf-1, an inhibitor of the Wnt signaling pathway, is induced by p53. Oncogene. 2000;19:1843 – 1848. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
