

Activation of peroxisome proliferator-activated receptor beta/delta inhibits lipopolysaccharide-induced cytokine production in adipocytes by lowering nuclear factor-kappaB activity via extracellular signal-related kinase 1/2
- PMID: 18443198
- PMCID: PMC2494695
- DOI: 10.2337/db08-0176
Activation of peroxisome proliferator-activated receptor beta/delta inhibits lipopolysaccharide-induced cytokine production in adipocytes by lowering nuclear factor-kappaB activity via extracellular signal-related kinase 1/2
Abstract
Objective: Chronic activation of the nuclear factor-kappaB (NF-kappaB) in white adipose tissue leads to increased production of pro-inflammatory cytokines, which are involved in the development of insulin resistance. It is presently unknown whether peroxisome proliferator-activated receptor (PPAR) beta/delta activation prevents inflammation in adipocytes.
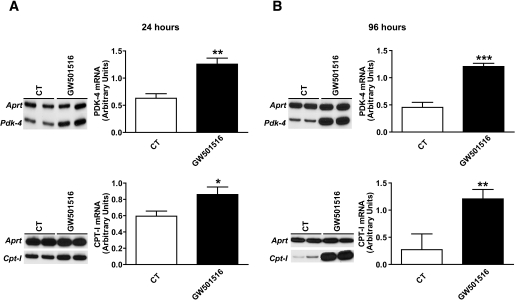
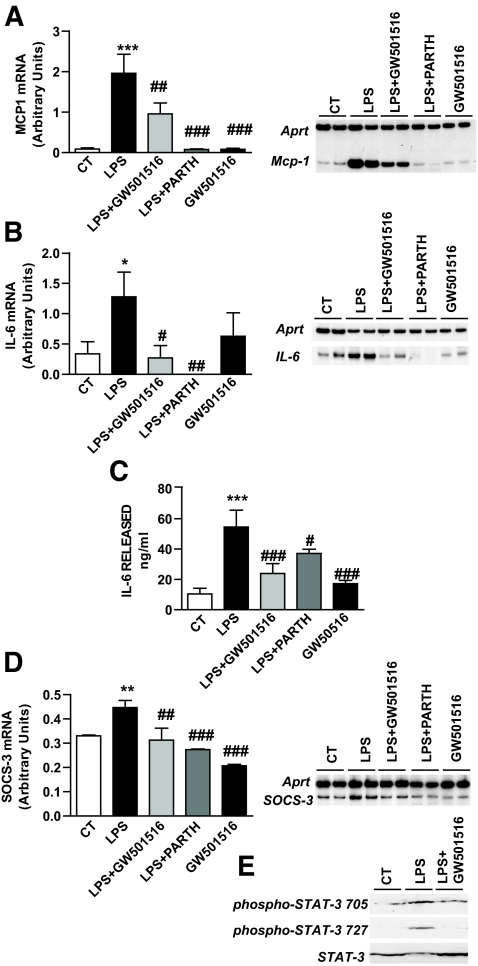
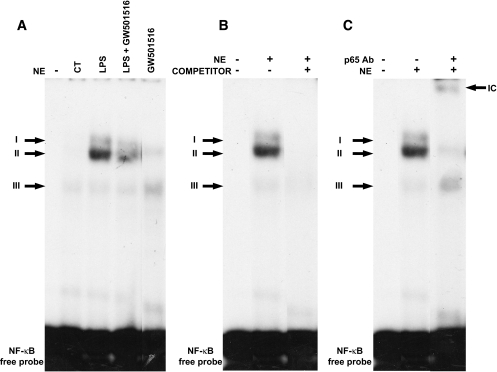
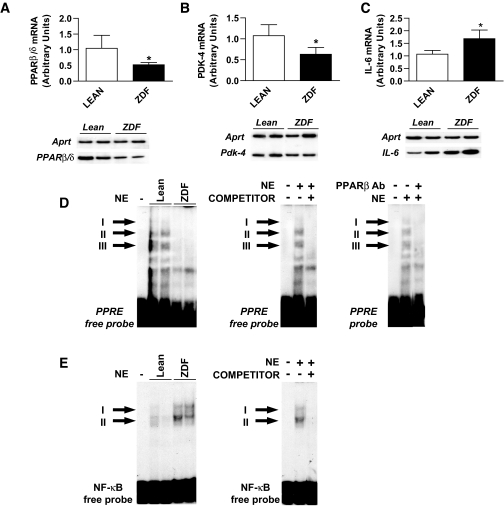
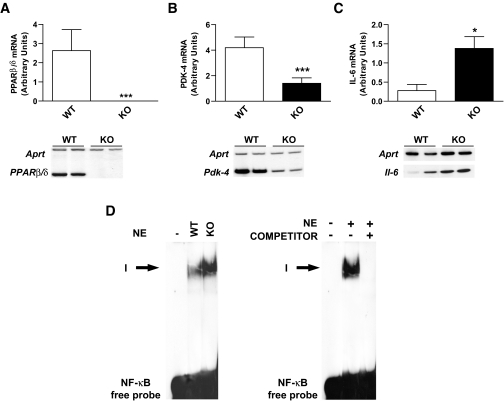
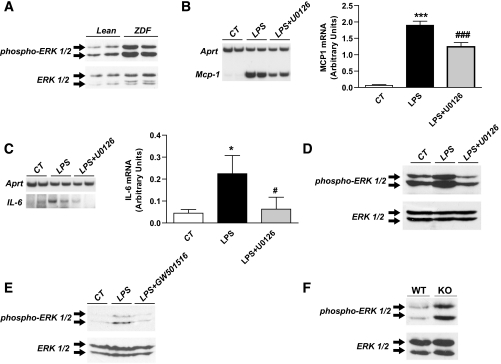
Research design and methods and results: First, we examined whether the PPARbeta/delta agonist GW501516 prevents lipopolysaccharide (LPS)-induced cytokine production in differentiated 3T3-L1 adipocytes. Treatment with GW501516 blocked LPS-induced IL-6 expression and secretion by adipocytes and the subsequent activation of the signal transducer and activator of transcription 3 (STAT3)-Suppressor of cytokine signaling 3 (SOCS3) pathway. This effect was associated with the capacity of GW501516 to impede LPS-induced NF-kappaB activation. Second, in in vivo studies, white adipose tissue from Zucker diabetic fatty (ZDF) rats, compared with that of lean rats, showed reduced PPARbeta/delta expression and PPAR DNA-binding activity, which was accompanied by enhanced IL-6 expression and NF-kappaB DNA-binding activity. Furthermore, IL-6 expression and NF-kappaB DNA-binding activity was higher in white adipose tissue from PPARbeta/delta-null mice than in wild-type mice. Because mitogen-activated protein kinase-extracellular signal-related kinase (ERK)1/2 (MEK1/2) is involved in LPS-induced NF-kappaB activation in adipocytes, we explored whether PPARbeta/delta prevented NF-kappaB activation by inhibiting this pathway. Interestingly, GW501516 prevented ERK1/2 phosphorylation by LPS. Furthermore, white adipose tissue from animal showing constitutively increased NF-kappaB activity, such as ZDF rats and PPARbeta/delta-null mice, also showed enhanced phospho-ERK1/2 levels.
Conclusions: These findings indicate that activation of PPARbeta/delta inhibits enhanced cytokine production in adipocytes by preventing NF-kappaB activation via ERK1/2, an effect that may help prevent insulin resistance.
Figures
References
-
- Hotamisligil GS, Shargill NS, Spiegelman BM: Adipose expression of tumor necrosis factor-alpha: direct role in obesity-linked insulin resistance. Science 259: 87–91, 1993 - PubMed
-
- Pickup JC, Mattock MB, Chusney GD, Burt D: NIDDM as a disease of the innate immune system: association of acute-phase reactants and interleukin-6 with metabolic syndrome X. Diabetologia 40: 1286–1292, 1997 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
