

Bone marrow-derived circulating endothelial precursors do not contribute to vascular endothelium and are not needed for tumor growth
- PMID: 18443294
- PMCID: PMC2365563
- DOI: 10.1073/pnas.0710516105
Bone marrow-derived circulating endothelial precursors do not contribute to vascular endothelium and are not needed for tumor growth
Abstract
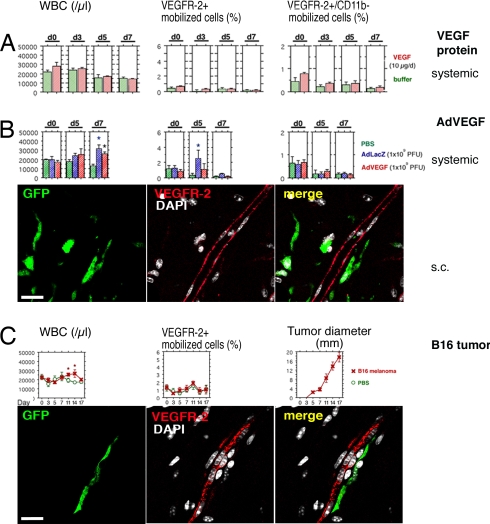
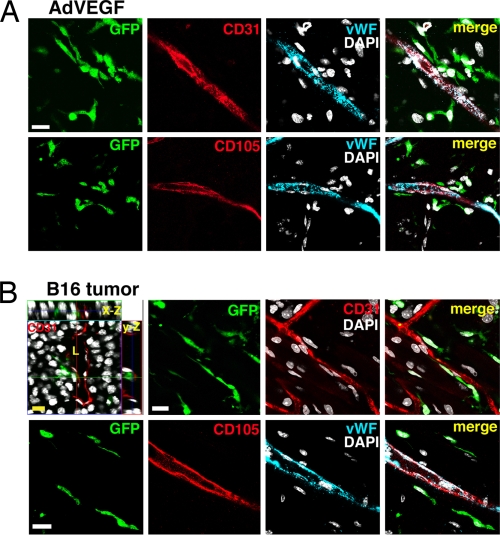
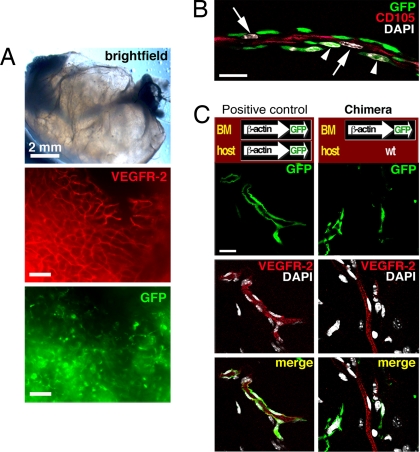
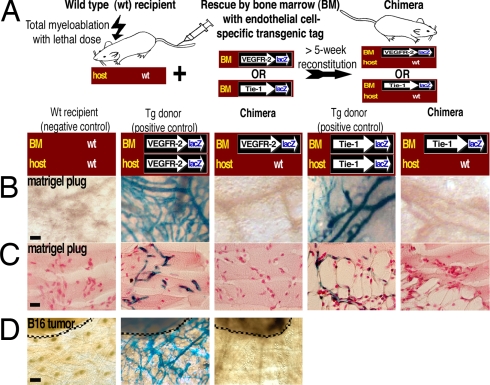
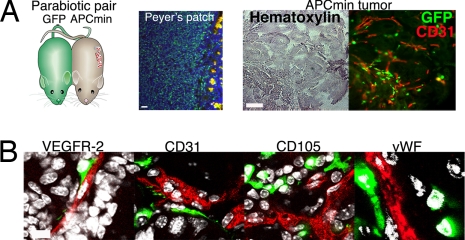
The mechanisms by which bone marrow (BM)-derived stem cells might contribute to angiogenesis and the origin of neovascular endothelial cells (ECs) are controversial. Neovascular ECs have been proposed to originate from VEGF receptor 2-expressing (VEGFR-2+) stem cells mobilized from the BM by VEGF or tumors, and it is thought that angiogenesis and tumor growth may depend on such endothelial precursors or progenitors. We studied the mobilization of BM cells to circulation by inoculating mice with VEGF polypeptides, adenoviral vectors expressing VEGF, or tumors. We induced angiogenesis by syngeneic melanomas, APCmin adenomas, adenoviral VEGF delivery, or matrigel plugs in four different genetically tagged universal or endothelial cell-specific chimeric mouse models, and subsequently analyzed the contribution of BM-derived cells to endothelium in a wide range of time points. To study the existence of circulating ECs in a nonmyeloablative setting, pairs of genetically marked parabiotic mice with a shared anastomosed circulatory system were created. We did not observe specific mobilization of VEGFR-2+ cells to circulation by VEGF or tumors. During angiogenesis, abundant BM-derived perivascular cells were recruited close to blood vessel wall ECs but did not form part of the endothelium. No circulation-derived vascular ECs were observed in the parabiosis experiments. Our results show that no BM-derived VEGFR-2+ or other EC precursors contribute to vascular endothelium and that cancer growth does not require BM-derived endothelial progenitors. Endothelial differentiation is not a typical in vivo function of normal BM-derived stem cells in adults, and it has to be an extremely rare event if it occurs at all.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Endothelial progenitor cells are cellular hubs essential for neoangiogenesis of certain aggressive adenocarcinomas and metastatic transition but not adenomas.Proc Natl Acad Sci U S A. 2008 Aug 26;105(34):E54; author reply E55. doi: 10.1073/pnas.0804876105. Epub 2008 Aug 20. Proc Natl Acad Sci U S A. 2008. PMID: 18715995 Free PMC article. No abstract available.
References
-
- Carmeliet P. Angiogenesis in life, disease and medicine. Nature. 2005;438:932–936. - PubMed
-
- Asahara T, et al. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275:964–967. - PubMed
-
- Takahashi T, et al. Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat Med. 1999;5:434–438. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
