

Widespread natural variation in murine natural killer T-cell number and function
- PMID: 18445005
- PMCID: PMC2669137
- DOI: 10.1111/j.1365-2567.2008.02846.x
Widespread natural variation in murine natural killer T-cell number and function
Abstract
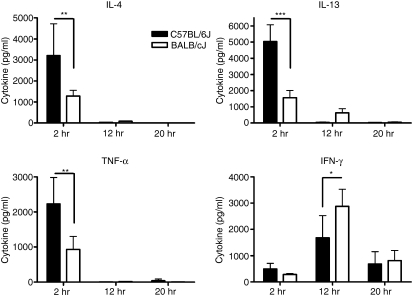
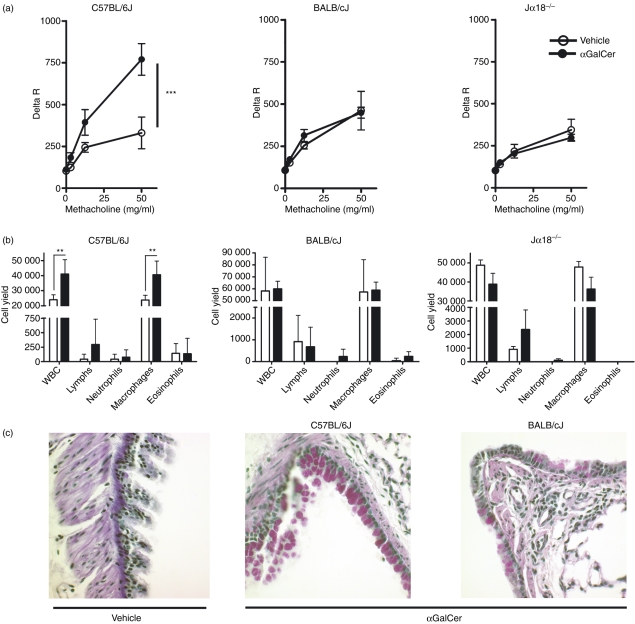
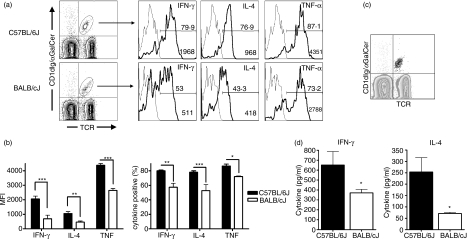
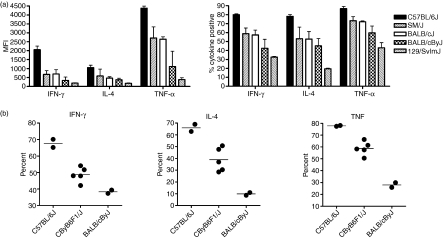
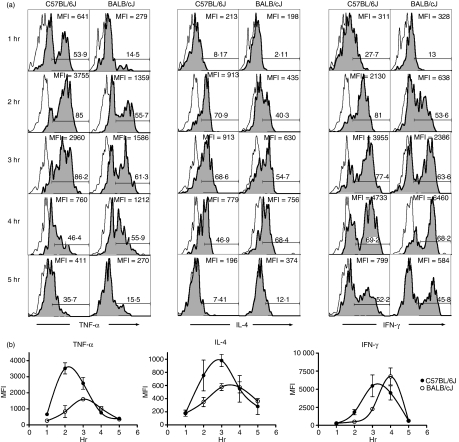
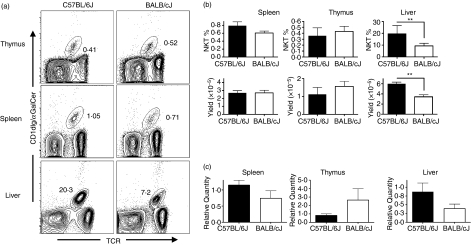
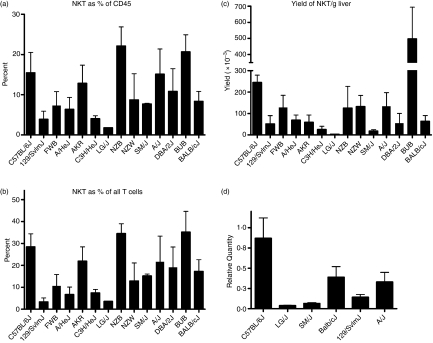
Natural killer T (NKT) cells comprise a novel T-lymphocyte subset that can influence a wide variety of immune responses through their ability to secrete large amounts of a variety of cytokines. Although variation in NKT-cell number and function has been extensively studied in autoimmune disease-prone mice, in which it has been linked to disease susceptibility, relatively little is known of the natural variation of NKT-cell number and function among normal inbred mouse strains. Here, we demonstrate strain-dependent variation in the susceptibility of C57BL/6J and BALB/cJ mice to NKT-mediated airway hyperreactivity, which correlated with significant increases in serum interleukin-4 (IL-4) and IL-13 elicited by the synthetic glycosphingolipid alpha-galactosylceramide. Examination of NKT-cell function revealed a significantly greater frequency of cytokine-producing NKT cells in C57BL/6J versus BALB/cJ mice as well as significant differences in the kinetics of NKT-cell cytokine production. Extension of this analysis to a panel of inbred mouse strains indicated that variability in NKT-cell cytokine production was widespread. Similarly, an examination of NKT-cell frequency revealed a significantly greater number of liver NKT cells in the C57BL/6J mice versus BALB/cJ mouse livers. Again, examination of a panel of inbred mouse strains revealed that liver NKT-cell numbers were quite variable, spanning over a 100-fold range. Taken together, these results demonstrate the presence of widespread natural variation in NKT-cell number and function among common inbred mouse strains, which may have implications for the examination of the influence of NKT cells in immune responses and disease pathogenesis among different genetic backgrounds.
Figures
References
-
- Kronenberg M. Toward an understanding of NKT cell biology: progress and paradoxes. Annu Rev Immunol. 2005;23:877–900. - PubMed
-
- Kawano T, Cui J, Koezuka Y, et al. CD1d-restricted and TCR-mediated activation of valpha14 NKT cells by glycosylceramides. Science. 1997;278:1626–9. - PubMed
-
- Brigl M, Bry L, Kent SC, Gumperz JE, Brenner MB. Mechanism of CD1d-restricted natural killer T cell activation during microbial infection. Nat Immunol. 2003;4:1230–7. - PubMed
-
- Paget C, Mallevaey T, Speak AO, et al. Activation of invariant NKT cells by toll-like receptor 9-stimulated dendritic cells requires type I interferon and charged glycosphingolipids. Immunity. 2007;27:597–609. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
