

Postsynaptic density-95 (PSD-95) and calcineurin control the sensitivity of N-methyl-D-aspartate receptors to calpain cleavage in cortical neurons
- PMID: 18445709
- PMCID: PMC2858625
- DOI: 10.1124/mol.108.046813
Postsynaptic density-95 (PSD-95) and calcineurin control the sensitivity of N-methyl-D-aspartate receptors to calpain cleavage in cortical neurons
Abstract
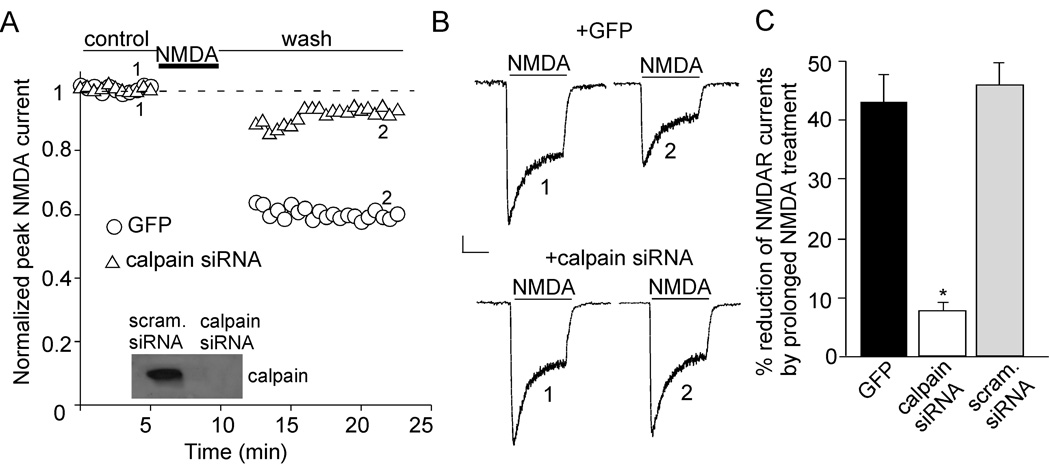
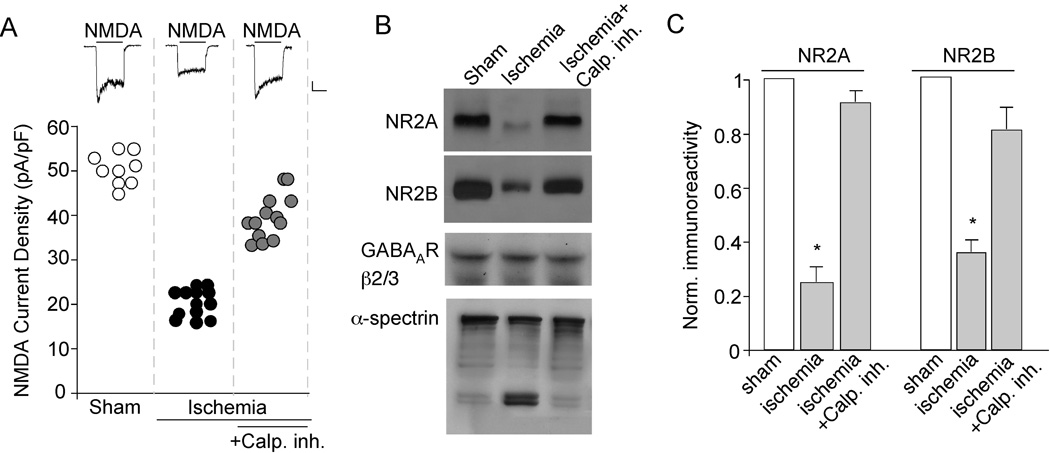
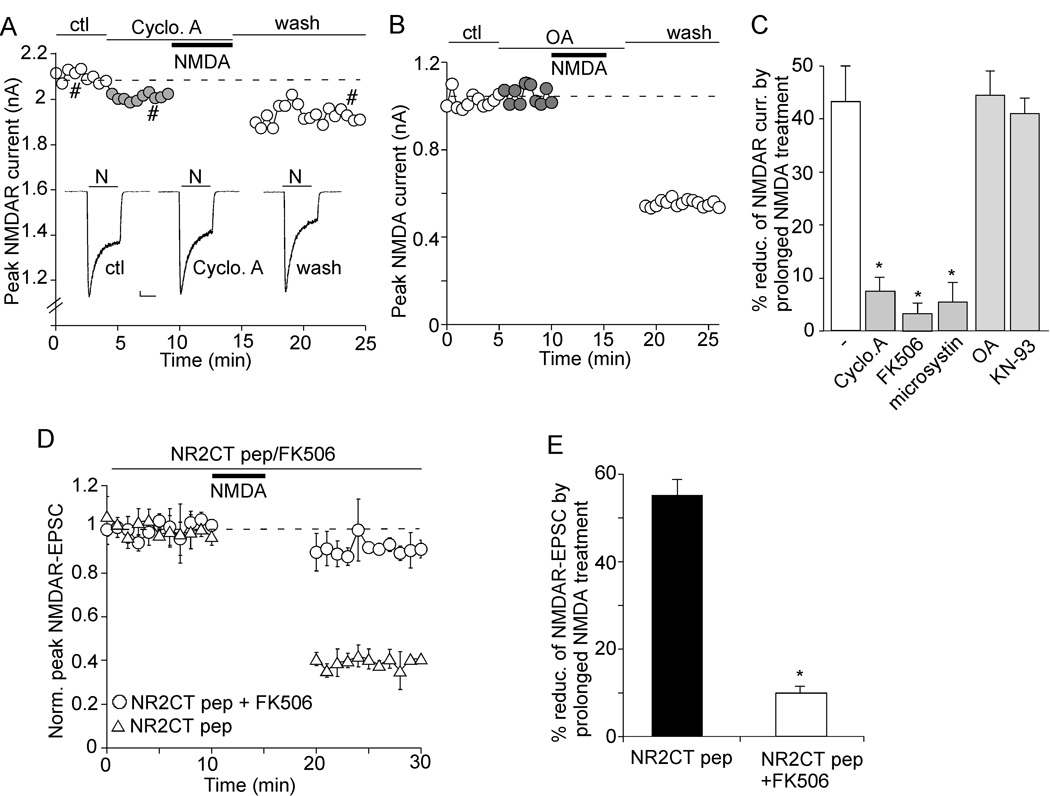
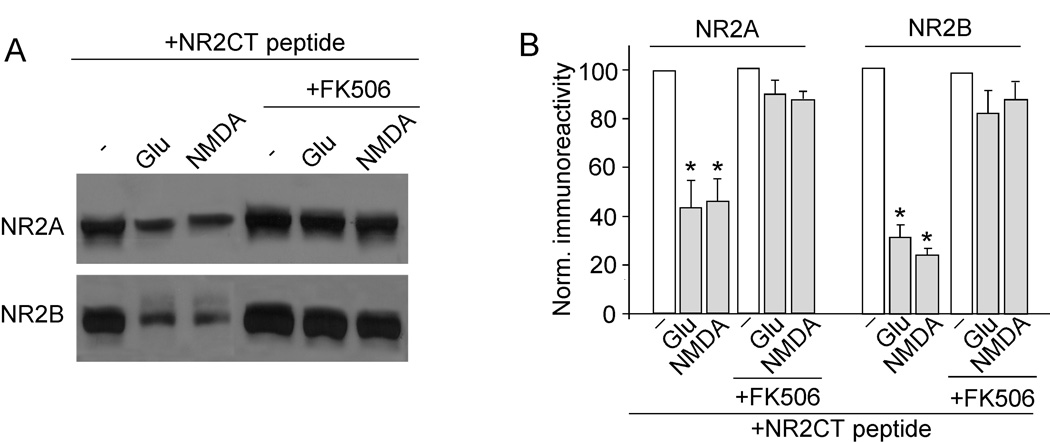
The N-methyl-D-aspartate receptor (NMDAR) is a Ca(2+)-permeable glutamate receptor mediating many neuronal functions under normal and pathological conditions. Ca(2+) influx via NMDARs activates diverse intracellular targets, including Ca(2+)-dependent protease calpain. Biochemical studies suggest that NR2A and NR2B subunits of NMDARs are substrates of calpain. Our physiological data showed that calpain, activated by prolonged NMDA treatment (100 microM, 5 min) of cultured cortical neurons, irreversibly decreased the whole-cell currents mediated by extrasynaptic NMDARs. Animals exposed to transient forebrain ischemia, a condition that activates calpain, exhibited the reduced NMDAR current density and the lower full-length NR2A/B level in a calpain-dependent manner. Disruption of the association between NMDARs and the scaffolding protein postsynaptic density (PSD)-95 facilitated the calpain regulation of synaptic NMDAR responses and NR2 cleavage in cortical slices, whereas inhibition of calcineurin activity blocked the calpain effect on NMDAR currents and NR2 cleavage. Calpain-cleaved NR2B subunits were removed from the cell surface. Moreover, cell viability assays showed that calpain, by targeting NMDARs, provided a negative feedback to dampen neuronal excitability in excitotoxic conditions. These data suggest that calpain activation suppresses NMDAR function via proteolytic cleavage of NR2 subunits in vitro and in vivo, and the susceptibility of NMDARs to calpain cleavage is controlled by PSD-95 and calcineurin.
Figures



References
-
- Aarts M, Liu Y, Liu L, Besshoh S, Arundine M, Gurd JW, Wang YT, Salter MW, Tymianski M. Treatment of ischemic brain damage by perturbing NMDA receptor- PSD-95 protein interactions. Science. 2002;298:846–850. - PubMed
-
- Abe K, Takeichi M. NMDA-receptor activation induces calpain-mediated beta-catenin cleavages for triggering gene expression. Neuron. 2007;53:387–397. - PubMed
-
- Adamec E, Beermann ML, Nixon RA. Calpain I activation in rat hippocampal neurons in culture is NMDA receptor selective and not essential for excitotoxic cell death. Brain Res Mol Brain Res. 1998;54:35–48. - PubMed
-
- Ankarcrona M, Dypbukt JM, Bonfoco E, Zhivotovsky B, Orrenius S, Lipton SA, Nicotera P. Glutamate-induced neuronal death: a succession of necrosis or apoptosis depending on mitochondrial function. Neuron. 1995;15:961–973. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
