

Extending the analysis of nicotinic receptor antagonists with the study of alpha6 nicotinic receptor subunit chimeras
- PMID: 18448138
- PMCID: PMC2494738
- DOI: 10.1016/j.neuropharm.2008.03.010
Extending the analysis of nicotinic receptor antagonists with the study of alpha6 nicotinic receptor subunit chimeras
Abstract
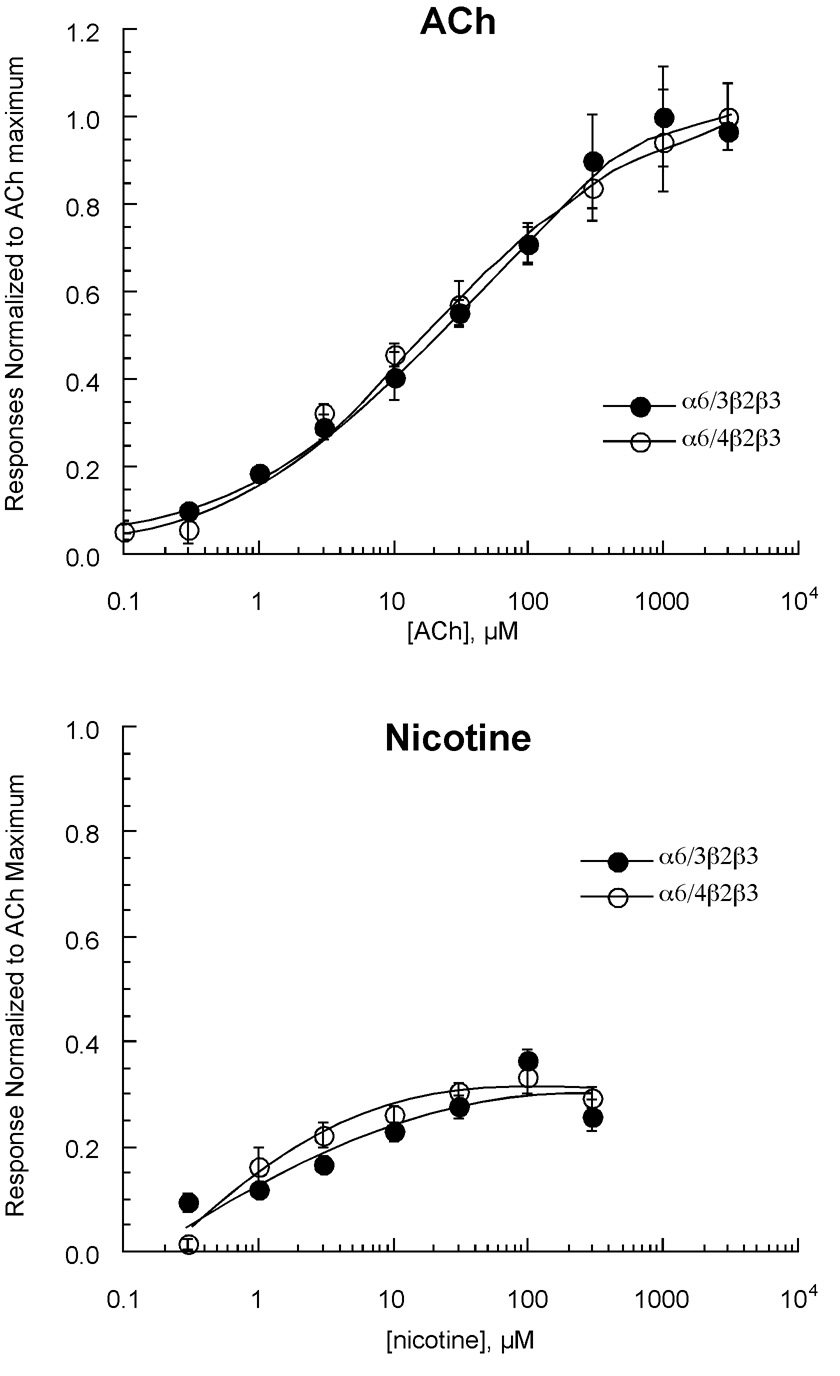
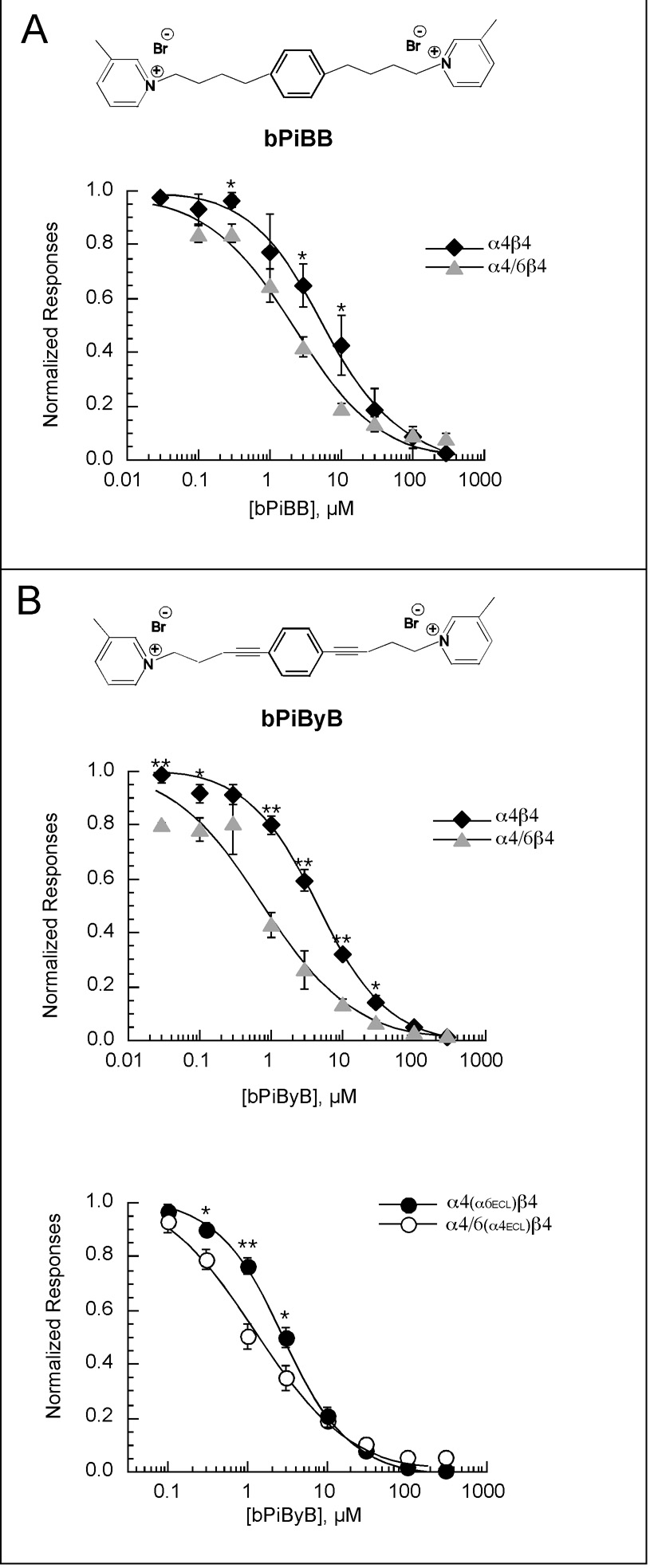
Heterologous expression systems have increased the feasibility of developing selective ligands to target nicotinic acetylcholine receptor (nAChR) subtypes. However, the alpha6 subunit, a component in nAChRs that mediates some of the reinforcing effects of nicotine, is not easily expressed in systems such as the Xenopus oocyte. Certain aspects of alpha6-containing receptor pharmacology have been studied by using chimeric subunits containing the alpha6 ligand-binding domain. However, these chimeras would not be sensitive to an alpha6-selective channel blocker; therefore we developed an alpha6 chimera (alpha4/6) that has the transmembrane and intracellular domains of alpha6 and the extracellular domain of alpha4. We examined the pharmacological properties of alpha4/6-containing receptors and other important nAChR subtypes, including alpha7, alpha4beta2, alpha4beta4, alpha3beta4, alpha3beta2, and alpha3beta2beta3, as well as receptors containing alpha6/3 and alpha6/4 chimeras. Our data show that the absence or presence of the beta4 subunit is an important factor for sensitivity to the ganglionic blocker mecamylamine, and that dihydro-beta-erythroidine is most effective on subtypes containing the alpha4 subunit extracellular domain. Receptors containing the alpha6/4 subunit are sensitive to alpha-conotoxin PIA, while receptors containing the reciprocal alpha4/6 chimera are insensitive. In experiments with novel antagonists of nicotine-evoked dopamine release, the alpha4/6 chimera indicated that structural rigidity was a key element of compounds that could result in selectivity for noncompetitive inhibition of alpha6-containing receptors. Our data extend the information available on prototypical nAChR antagonists, and establish the alpha4/6 chimera as a useful new tool for screening drugs as selective nAChR antagonists.
Figures








Similar articles
-
A novel α4/7-conotoxin LvIA from Conus lividus that selectively blocks α3β2 vs. α6/α3β2β3 nicotinic acetylcholine receptors.FASEB J. 2014 Apr;28(4):1842-53. doi: 10.1096/fj.13-244103. Epub 2014 Jan 7. FASEB J. 2014. PMID: 24398291 Free PMC article.
-
A role for α4(non-α6)* nicotinic acetylcholine receptors in motor behavior.Neuropharmacology. 2013 Oct;73:19-30. doi: 10.1016/j.neuropharm.2013.05.001. Epub 2013 May 17. Neuropharmacology. 2013. PMID: 23688922 Free PMC article.
-
Pharmacological and functional comparisons of α6/α3β2β3-nAChRs and α4β2-nAChRs heterologously expressed in the human epithelial SH-EP1 cell line.Acta Pharmacol Sin. 2018 Oct;39(10):1571-1581. doi: 10.1038/aps.2017.209. Epub 2018 May 24. Acta Pharmacol Sin. 2018. PMID: 29795357 Free PMC article.
-
Mysterious alpha6-containing nAChRs: function, pharmacology, and pathophysiology.Acta Pharmacol Sin. 2009 Jun;30(6):740-51. doi: 10.1038/aps.2009.63. Acta Pharmacol Sin. 2009. PMID: 19498417 Free PMC article. Review.
-
Striatal alpha6* nicotinic acetylcholine receptors: potential targets for Parkinson's disease therapy.J Pharmacol Exp Ther. 2006 Feb;316(2):481-9. doi: 10.1124/jpet.105.094375. Epub 2005 Oct 6. J Pharmacol Exp Ther. 2006. PMID: 16210393 Review.
Cited by
-
Evaluation of structurally diverse neuronal nicotinic receptor ligands for selectivity at the alpha6( *) subtype.Bioorg Med Chem Lett. 2009 Aug 1;19(15):4359-63. doi: 10.1016/j.bmcl.2009.05.085. Epub 2009 May 27. Bioorg Med Chem Lett. 2009. PMID: 19560354 Free PMC article.
-
Molecular Determinants of Species Specificity of α-Conotoxin TxIB towards Rat and Human α6/α3β4 Nicotinic Acetylcholine Receptors.Int J Mol Sci. 2023 May 11;24(10):8618. doi: 10.3390/ijms24108618. Int J Mol Sci. 2023. PMID: 37239959 Free PMC article.
-
The contribution of α4β2 and non-α4β2 nicotinic acetylcholine receptors to the discriminative stimulus effects of nicotine and varenicline in mice.Psychopharmacology (Berl). 2017 Mar;234(5):781-792. doi: 10.1007/s00213-016-4514-4. Epub 2016 Dec 27. Psychopharmacology (Berl). 2017. PMID: 28028600 Free PMC article.
-
The Concise Guide to PHARMACOLOGY 2013/14: ligand-gated ion channels.Br J Pharmacol. 2013 Dec;170(8):1582-606. doi: 10.1111/bph.12446. Br J Pharmacol. 2013. PMID: 24528238 Free PMC article.
-
Comparison of effects produced by nicotine and the α4β2-selective agonist 5-I-A-85380 on intracranial self-stimulation in rats.Exp Clin Psychopharmacol. 2016 Feb;24(1):65-75. doi: 10.1037/pha0000055. Exp Clin Psychopharmacol. 2016. PMID: 26461167 Free PMC article.
References
-
- Ayers JT, Dwoskin LP, Deaciuc AG, Grinevich VP, Zhu J, Crooks PA. bis-Azaaromatic quaternary ammonium analogues: ligands for alpha4beta2* and alpha7* subtypes of neuronal nicotinic receptors. Bioorganic Medicinal Chemistry Letters. 2002;12(21):3067–3071. - PubMed
-
- Azam L, McIntosh JM. Characterization of nicotinic acetylcholine receptors that modulate nicotine-evoked [3H]norepinephrine release from mouse hippocampal synaptosomes. Molecular Pharmacology. 2006;70(3):967–976. - PubMed
-
- Bouzat C, Gumilar F, Spitzmaul G, Wang HL, Rayes D, Hansen SB, Taylor P, Sine SM. Coupling of agonist binding to channel gating in an ACh-binding protein linked to an ion channel. Nature. 2004;430(7002):896–900. - PubMed
-
- Charpantier E, Barneoud P, Moser P, Besnard F, Sgard F. Nicotinic acetylcholine subunit mRNA expression in dopaminergic neurons of the rat substantia nigra and ventral tegmental area. Neuroreport. 1998;9(13):3097–3101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
