

Bam32: a novel mediator of Erk activation in T cells
- PMID: 18448454
- PMCID: PMC2680712
- DOI: 10.1093/intimm/dxn039
Bam32: a novel mediator of Erk activation in T cells
Abstract
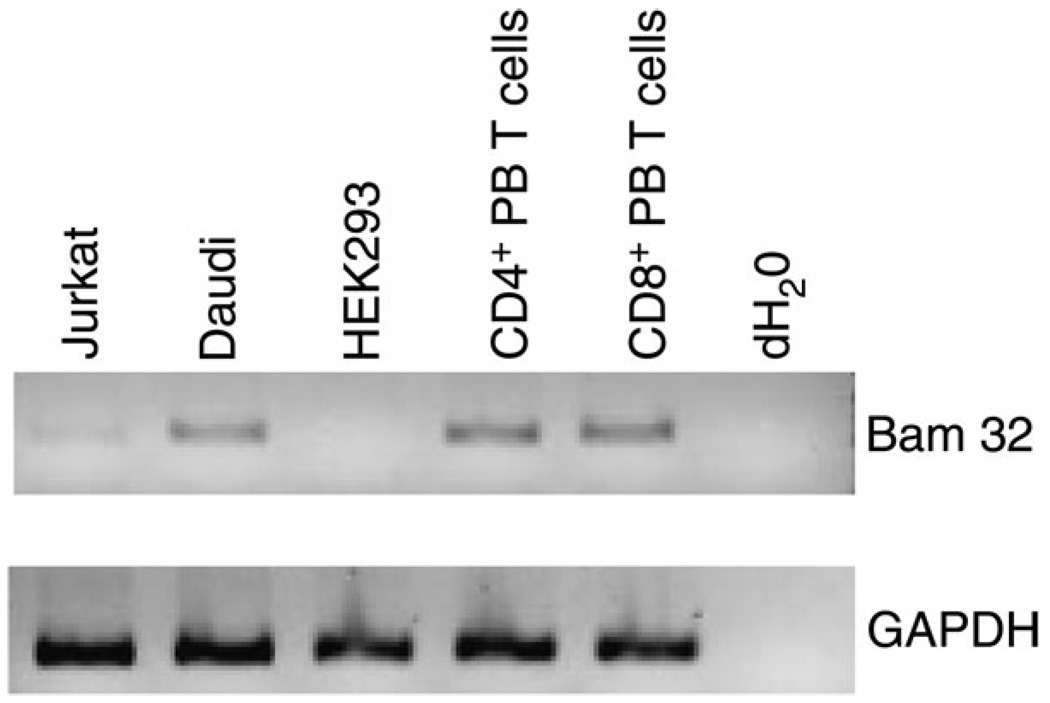
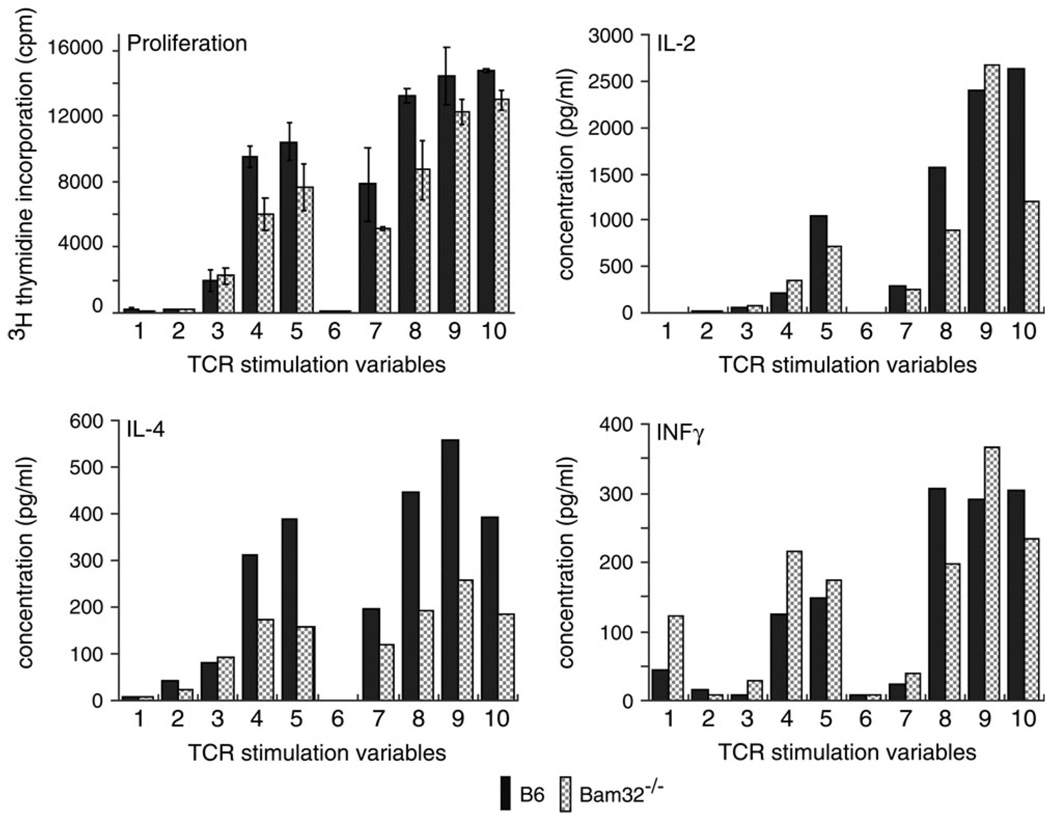
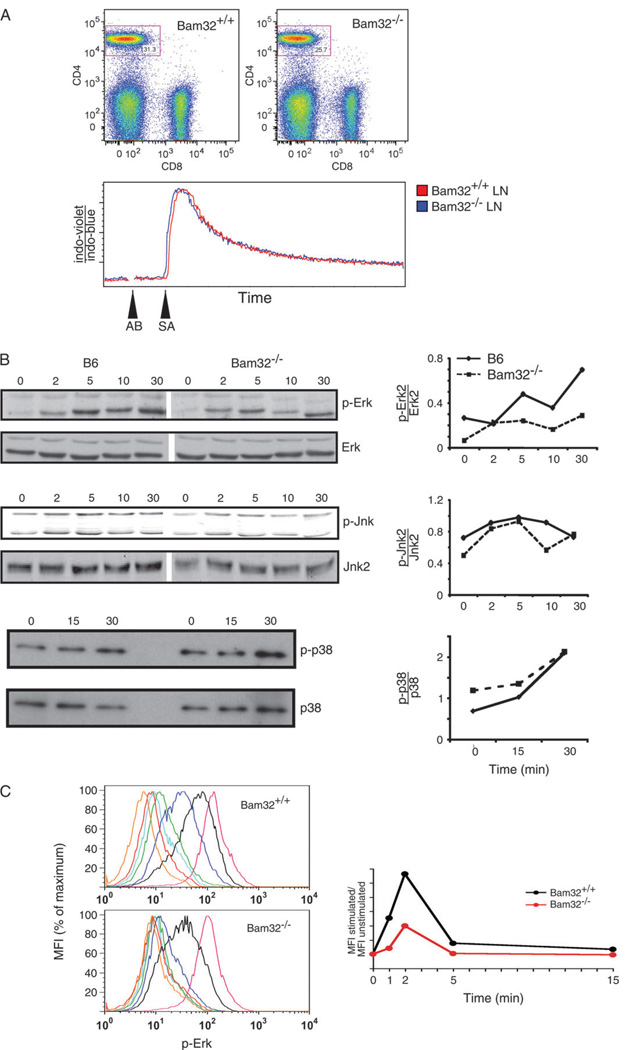
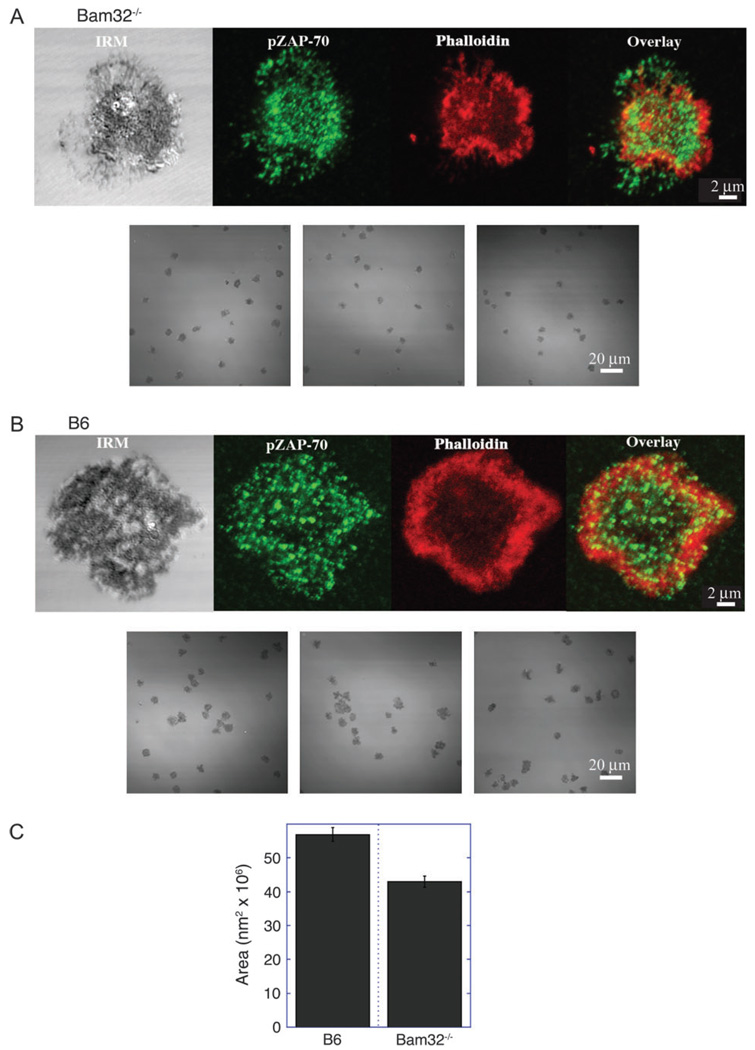
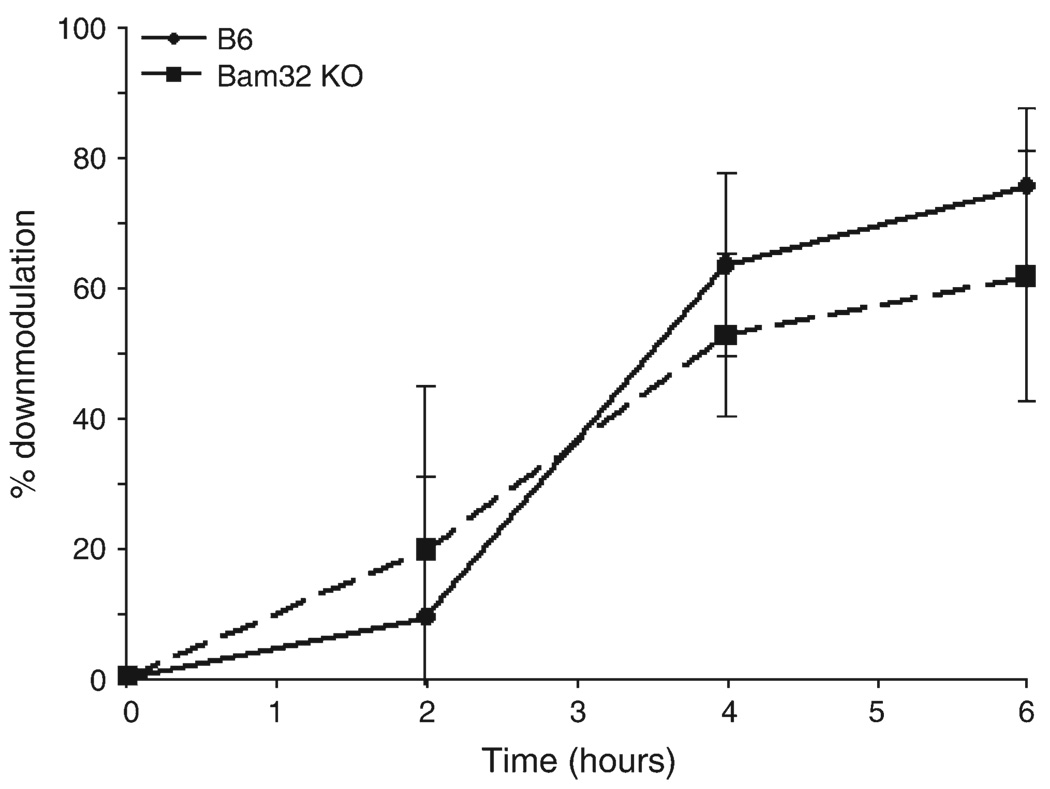
Bam32 (B lymphocyte adapter molecule of 32 kDa) is an adapter protein expressed in some hematopoietic cells including B and T lymphocytes. It was previously shown that Bam32-deficient mice have defects in various aspects of B cell activation including B cell receptor (BCR)-induced Erk activation, BCR-induced proliferation and T-independent antibody responses. In this study, we have examined the role of Bam32 in T cell activation using Bam32-deficient mice. By comparing CD4(+) T cells from lymph nodes of wild-type and Bam32-deficient mice, we found that Bam32 was required for optimal TCR-induced Erk activation, cytokine production, proliferation and actin-mediated spreading of CD4(+) T cells. These results indicate a novel pathway to Erk activation in T cells involving the adapter protein Bam32.
Figures
References
-
- Allam A, Marshall AJ. Role of the adaptor proteins Bam32, TAPP1 and TAPP2 in lymphocyte activation. Immunol. Lett. 2005;97:7. - PubMed
-
- Han A, Saijo K, Mecklenbrauker I, Tarakhovsky A, Nussenzweig MC. Bam32 links the B cell receptor to ERK and JNK and mediates B cell proliferation but not survival. Immunity. 2003;19:621. - PubMed
-
- Rao VR, Corradetti MN, Chen J, et al. Expression cloning of protein targets for 3-phosphorylated phosphoinositides. J. Biol. Chem. 1999;274:37893. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
