

Biogenesis of extracellular microfibrils: Multimerization of the fibrillin-1 C terminus into bead-like structures enables self-assembly
- PMID: 18448684
- PMCID: PMC2373353
- DOI: 10.1073/pnas.0706335105
Biogenesis of extracellular microfibrils: Multimerization of the fibrillin-1 C terminus into bead-like structures enables self-assembly
Abstract
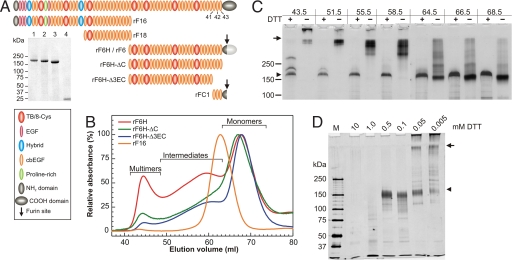
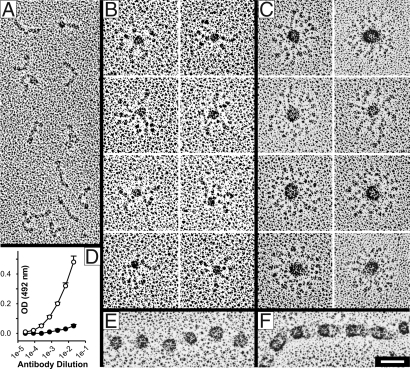
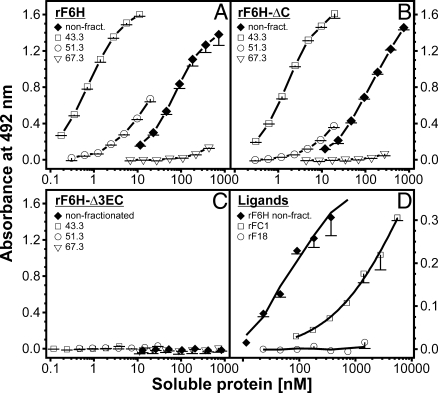
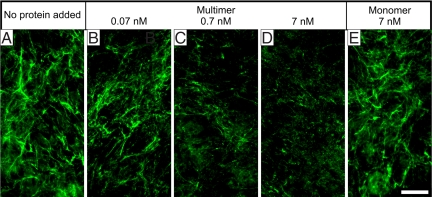
Microfibrils are essential elements in elastic and nonelastic tissues contributing to homeostasis and growth factor regulation. Fibrillins form the core of these multicomponent assemblies. Various human genetic disorders, the fibrillinopathies, arise from mutations in fibrillins and are frequently associated with aberrant microfibril assembly. These disorders include Marfan syndrome, Weill-Marchesani syndrome, Beals syndrome, and others. Although homotypic and heterotypic fibrillin self-interactions are considered to provide critical initial steps, the detailed mechanisms for microfibril assembly are unknown. We show here that the C-terminal recombinant half of fibrillin-1 assembles into disulfide-bonded multimeric globular structures with peripheral arms and a dense core. These globules are similar to the beaded structures observed in microfibrils isolated from tissues. Only these C-terminal fibrillin-1 multimers interacted strongly with the fibrillin-1 N terminus, whereas the monomers showed very little self-interaction activity. The multimers strongly inhibited microfibril formation in cell culture, providing evidence that these recombinant assemblies can also interact with endogenous fibrillin-1. The C-terminal self-interaction site was fine-mapped to the last three calcium-binding EGF domains in fibrillin-1. These results suggest a new mechanism for microfibril formation where fibrillin-1 first oligomerizes via its C terminus before the partially or fully assembled bead-like structures can further interact with other beads via the fibrillin-1 N termini.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Hubmacher D, Tiedemann K, Reinhardt DP. Fibrillins: From biogenesis of microfibrils to signaling functions. Curr Top Dev Biol. 2006;75:93–123. - PubMed
-
- Kielty CM. Elastic fibres in health and disease. Expert Rev Mol Med. 2006;8:1–23. - PubMed
-
- Henderson M, Polewski R, Fanning JC, Gibson MA. Microfibril-associated glycoprotein-1 (MAGP-1) is specifically located on the beads of the beaded-filament structure for fibrillin-containing microfibrils as visualized by the rotary shadowing technique. J Histochem Cytochem. 1996;44:1389–1397. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
