

SDF-1/CXCR4 and VLA-4 interaction regulates homing in Waldenstrom macroglobulinemia
- PMID: 18448868
- PMCID: PMC2435685
- DOI: 10.1182/blood-2007-12-129395
SDF-1/CXCR4 and VLA-4 interaction regulates homing in Waldenstrom macroglobulinemia
Abstract
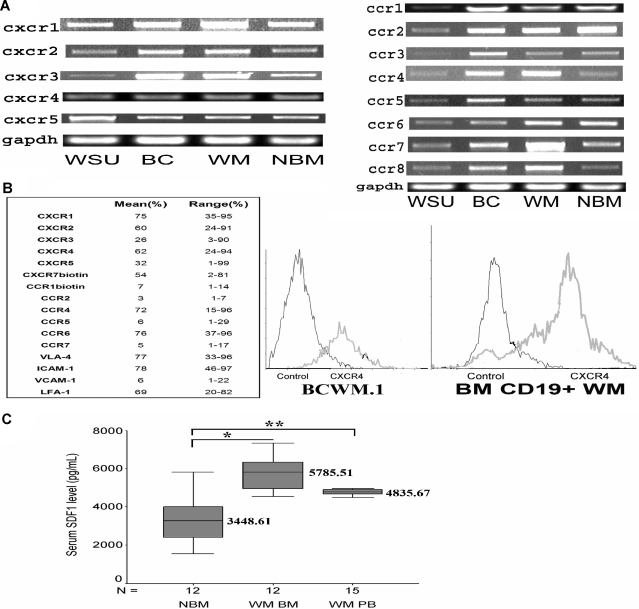
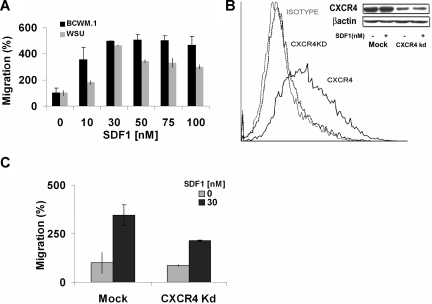
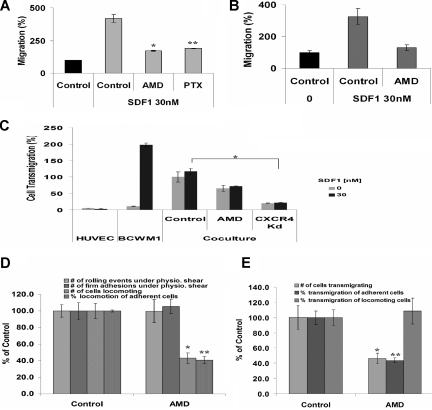
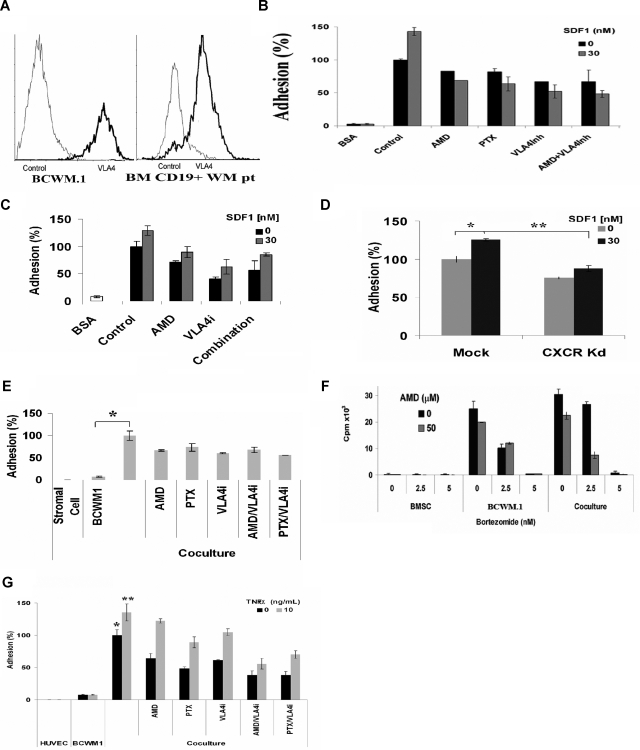
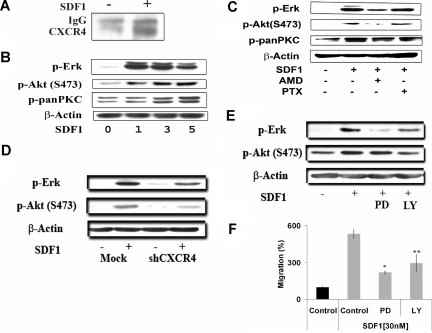
Waldenstrom macroglobulinemia (WM) is characterized by widespread involvement of the bone marrow at the time of diagnosis, implying continuous homing of WM cells into the marrow. The mechanisms by which trafficking of the malignant cells into the bone marrow has not been previously elucidated. In this study, we show that WM cells express high levels of chemokine and adhesion receptors, including CXCR4 and VLA-4. We showed that CXCR4 was essential for the migration and trans-endothelial migration of WM cells under static and dynamic shear flow conditions, with significant inhibition of migration using CXCR4 knockdown or the CXCR4 inhibitor AMD3100. Similarly, CXCR4 or VLA-4 inhibition led to significant inhibition of adhesion to fibronectin, stromal cells, and endothelial cells. Decreased adhesion of WM cells to stromal cells by AMD3100 led to increased sensitivity of these cells to cytotoxicity by bortezomib. To further investigate the mechanisms of CXCR4-dependent adhesion, we showed that CXCR4 and VLA-4 directly interact in response to SDF-1, we further investigated downstream signaling pathways regulating migration and adhesion in WM. Together, these studies demonstrate that the CXCR4/SDF-1 axis interacts with VLA-4 in regulating migration and adhesion of WM cells in the bone marrow microenvironment.
Figures
References
-
- Ghobrial IM, Gertz MA, Fonseca R. Waldenstrom macroglobulinaemia. Lancet Oncol. 2003;4:679–685. - PubMed
-
- Vijay A, Gertz MA. Waldenstrom macroglobulinemia. Blood. 2007;109:5096–5103. - PubMed
-
- Ghobrial IM, Fonseca R, Gertz MA, et al. Prognostic model for disease-specific and overall mortality in newly diagnosed symptomatic patients with Waldenstrom macroglobulinaemia. Br J Haematol. 2006;133:158–164. - PubMed
-
- Baggiolini M. Chemokines and leukocyte traffic. Nature. 1998;392:565–568. - PubMed
-
- Murphy PM. Chemokines and the molecular basis of cancer metastasis. N Engl J Med. 2001;345:833–835. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
