

Erasing the methyl mark: histone demethylases at the center of cellular differentiation and disease
- PMID: 18451103
- PMCID: PMC2732404
- DOI: 10.1101/gad.1652908
Erasing the methyl mark: histone demethylases at the center of cellular differentiation and disease
Abstract
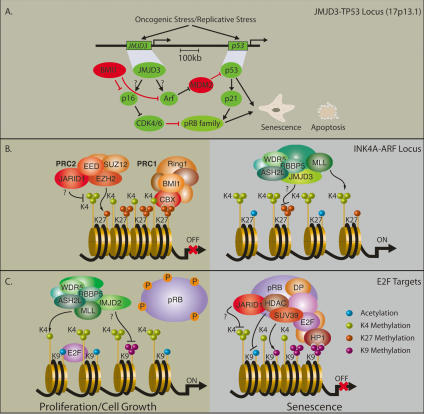
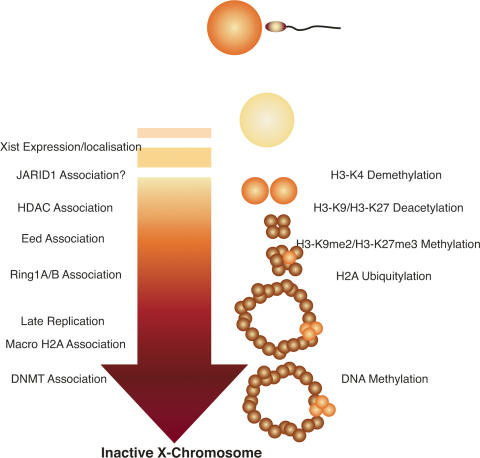
The enzymes catalyzing lysine and arginine methylation of histones are essential for maintaining transcriptional programs and determining cell fate and identity. Until recently, histone methylation was regarded irreversible. However, within the last few years, several families of histone demethylases erasing methyl marks associated with gene repression or activation have been identified, underscoring the plasticity and dynamic nature of histone methylation. Recent discoveries have revealed that histone demethylases take part in large multiprotein complexes synergizing with histone deacetylases, histone methyltransferases, and nuclear receptors to control developmental and transcriptional programs. Here we review the emerging biochemical and biological functions of the histone demethylases and discuss their potential involvement in human diseases, including cancer.
Figures
References
-
- Agger K., Cloos P.A., Christensen J., Pasini D., Rose S., Rappsilber J., Issaeva I., Canaani E., Salcini A.E., Helin K. UTX and JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. Nature. 2007;449:731–734. - PubMed
-
- Akasaka T., Takahashi N., Suzuki M., Koseki H., Bodmer R., Koga H. MBLR, a new RING finger protein resembling mammalian Polycomb gene products, is regulated by cell cycle-dependent phosphorylation. Genes Cells. 2002;7:835–850. - PubMed
-
- Allis C.D., Berger S.L., Cote J., Dent S., Jenuwien T., Kouzarides T., Pillus L., Reinberg D., Shi Y., Shiekhattar R., et al. New nomenclature for chromatin-modifying enzymes. Cell. 2007;131:633–636. - PubMed
-
- Bannister A.J., Zegerman P., Partridge J.F., Miska E.A., Thomas J.O., Allshire R.C., Kouzarides T. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature. 2001;410:120–124. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources