

The role of the chromatin remodeler Mi-2beta in hematopoietic stem cell self-renewal and multilineage differentiation
- PMID: 18451107
- PMCID: PMC2335314
- DOI: 10.1101/gad.1642808
The role of the chromatin remodeler Mi-2beta in hematopoietic stem cell self-renewal and multilineage differentiation
Abstract
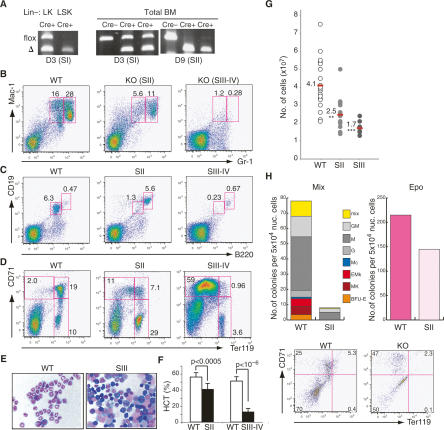
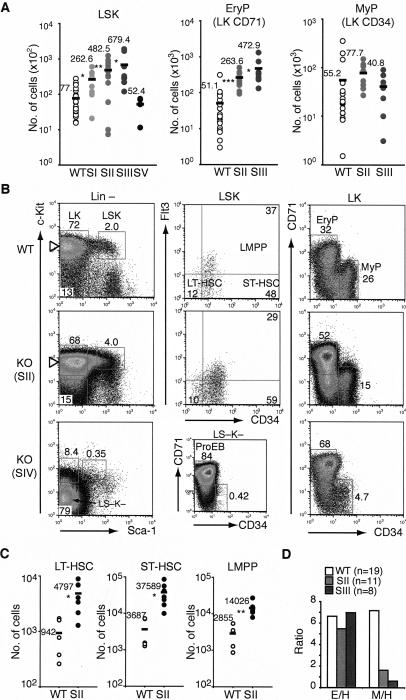
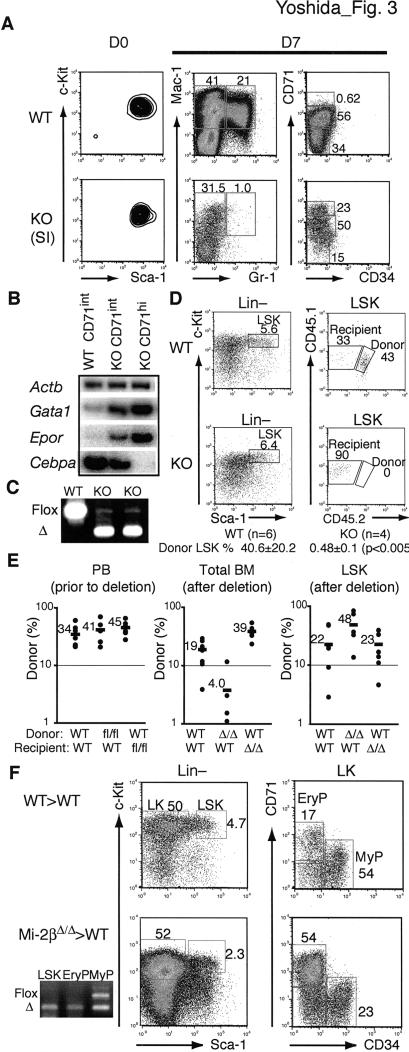
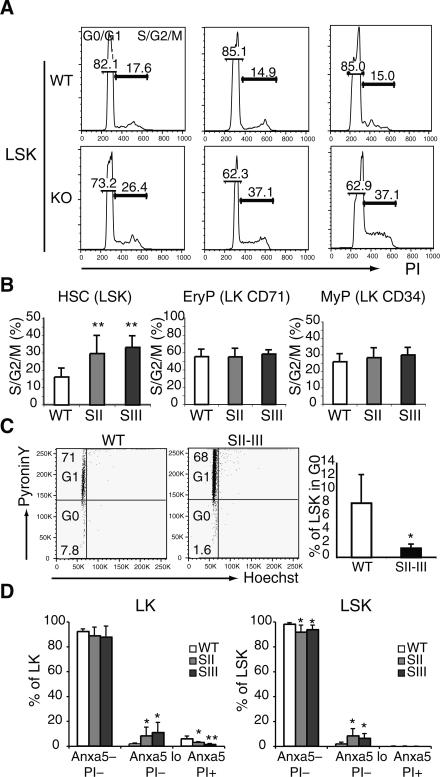
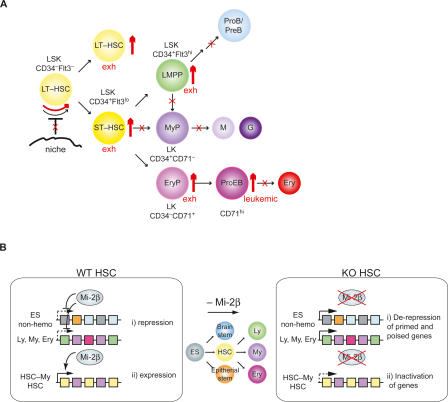
The ability of somatic stem cells to self-renew and differentiate into downstream lineages is dependent on specialized chromatin environments that keep stem cell-specific genes active and key differentiation factors repressed but poised for activation. The epigenetic factors that provide this type of regulation remain ill-defined. Here we provide the first evidence that the SNF2-like ATPase Mi-2beta of the Nucleosome Remodeling Deacetylase (NuRD) complex is required for maintenance of and multilineage differentiation in the early hematopoietic hierarchy. Shortly after conditional inactivation of Mi-2beta, there is an increase in cycling and a decrease in quiescence in an HSC (hematopoietic stem cell)-enriched bone marrow population. These cycling mutant cells readily differentiate into the erythroid lineage but not into the myeloid and lymphoid lineages. Together, these effects result in an initial expansion of mutant HSC and erythroid progenitors that are later depleted as more differentiated proerythroblasts accumulate at hematopoietic sites exhibiting features of erythroid leukemia. Examination of gene expression in the mutant HSC reveals changes in the expression of genes associated with self-renewal and lineage priming and a pivotal role of Mi-2beta in their regulation. Thus, Mi-2beta provides the hematopoietic system with immune cell capabilities as well as with an extensive regenerative capacity.
Figures
Similar articles
-
Chromatin restriction by the nucleosome remodeler Mi-2β and functional interplay with lineage-specific transcription regulators control B-cell differentiation.Genes Dev. 2019 Jul 1;33(13-14):763-781. doi: 10.1101/gad.321901.118. Epub 2019 May 23. Genes Dev. 2019. PMID: 31123064 Free PMC article.
-
Flk2/Flt3 promotes both myeloid and lymphoid development by expanding non-self-renewing multipotent hematopoietic progenitor cells.Exp Hematol. 2014 Mar;42(3):218-229.e4. doi: 10.1016/j.exphem.2013.11.013. Epub 2013 Dec 11. Exp Hematol. 2014. PMID: 24333663 Free PMC article.
-
Reduced lymphoid lineage priming promotes human hematopoietic stem cell expansion.Cell Stem Cell. 2014 Jan 2;14(1):94-106. doi: 10.1016/j.stem.2013.11.021. Cell Stem Cell. 2014. PMID: 24388174
-
Concise review: Multidimensional regulation of the hematopoietic stem cell state.Stem Cells. 2012 Jan;30(1):82-8. doi: 10.1002/stem.776. Stem Cells. 2012. PMID: 22083966 Review.
-
The Mi-2/NuRD complex: a critical epigenetic regulator of hematopoietic development, differentiation and cancer.Epigenetics. 2009 Nov 16;4(8):532-6. doi: 10.4161/epi.4.8.10108. Epub 2009 Nov 16. Epigenetics. 2009. PMID: 19923891 Review.
Cited by
-
The Chromatin Remodeler LET-418/Mi2 is Required Cell Non-Autonomously for the Post-Embryonic Development of Caenorhabditis elegans.J Dev Biol. 2018 Dec 24;7(1):1. doi: 10.3390/jdb7010001. J Dev Biol. 2018. PMID: 30586943 Free PMC article.
-
GFI1 tethers the NuRD complex to open and transcriptionally active chromatin in myeloid progenitors.Commun Biol. 2021 Dec 2;4(1):1356. doi: 10.1038/s42003-021-02889-2. Commun Biol. 2021. PMID: 34857890 Free PMC article.
-
ATP-dependent chromatin remodeling complexes as novel targets for cancer therapy.Adv Cancer Res. 2014;121:183-233. doi: 10.1016/B978-0-12-800249-0.00005-6. Adv Cancer Res. 2014. PMID: 24889532 Free PMC article. Review.
-
Roles of p53 in various biological aspects of hematopoietic stem cells.J Biomed Biotechnol. 2012;2012:903435. doi: 10.1155/2012/903435. Epub 2012 Jun 20. J Biomed Biotechnol. 2012. PMID: 22778557 Free PMC article. Review.
-
Genomic transcription factor binding site selection is edited by the chromatin remodeling factor CHD4.Nucleic Acids Res. 2024 Apr 24;52(7):3607-3622. doi: 10.1093/nar/gkae025. Nucleic Acids Res. 2024. PMID: 38281186 Free PMC article.
References
-
- Adams G.B., Chabner K.T., Alley I.R., Olson D.P., Szczepiorkowski Z.M., Poznansky M.C., Kos C.H., Pollak M.R., Brown E.M., Scadden D.T. Stem cell engraftment at the endosteal niche is specified by the calcium-sensing receptor. Nature. 2006;439:599–603. - PubMed
-
- Adolfsson J., Mansson R., Buza-Vidas N., Hultquist A., Liuba K., Jensen C.T., Bryder D., Yang L., Borge O.J., Thoren L.A., et al. Identification of Flt3+ lympho-myeloid stem cells lacking erythro-megakaryocytic potential a revised road map for adult blood lineage commitment. Cell. 2005;121:295–306. - PubMed
-
- Akashi K., Traver D., Miyamoto T., Weissman I.L. A clonogenic common myeloid progenitor that gives rise to all myeloid lineages. Nature. 2000;404:193–197. - PubMed
-
- Arai F., Hirao A., Ohmura M., Sato H., Matsuoka S., Takubo K., Ito K., Koh G.Y., Suda T. Tie2/angiopoietin-1 signaling regulates hematopoietic stem cell quiescence in the bone marrow niche. Cell. 2004;118:149–161. - PubMed
-
- Bernstein B.E., Mikkelsen T.S., Xie X., Kamal M., Huebert D.J., Cuff J., Fry B., Meissner A., Wernig M., Plath K., et al. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell. 2006;125:315–326. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases