

CD151 accelerates breast cancer by regulating alpha 6 integrin function, signaling, and molecular organization
- PMID: 18451146
- PMCID: PMC4764302
- DOI: 10.1158/0008-5472.CAN-07-2949
CD151 accelerates breast cancer by regulating alpha 6 integrin function, signaling, and molecular organization
Abstract
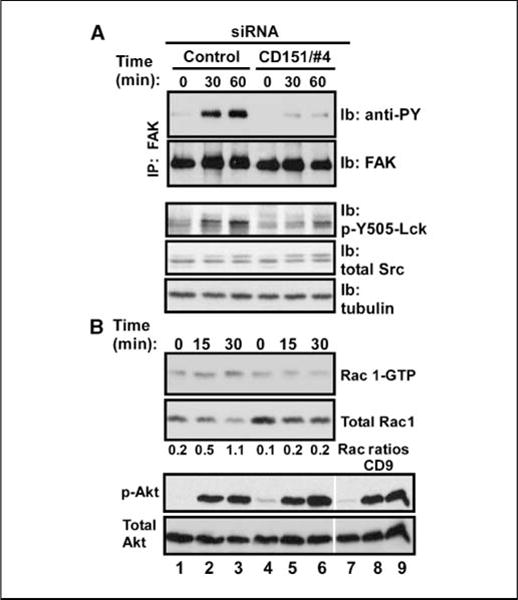
CD151, a master regulator of laminin-binding integrins (alpha(6)beta(4), alpha(6)beta(1), and alpha(3)beta(1)), assembles these integrins into complexes called tetraspanin-enriched microdomains. CD151 protein expression is elevated in 31% of human breast cancers and is even more elevated in high-grade (40%) and estrogen receptor-negative (45%) subtypes. The latter includes triple-negative (estrogen receptor, progesterone receptor, and HER2 negative) basal-like tumors. CD151 ablation markedly reduced basal-like mammary cell migration, invasion, spreading, and signaling (through FAK, Rac1, and lck) while disrupting epidermal growth factor receptor (EGFR)-alpha(6) integrin collaboration. Underlying these defects, CD151 ablation redistributed alpha(6)beta(4) integrins subcellularly and severed molecular links between integrins and tetraspanin-enriched microdomains. In a prototypical basal-like mammary tumor line, CD151 ablation notably delayed tumor progression in ectopic and orthotopic xenograft models. These results (a) establish that CD151-alpha(6) integrin complexes play a functional role in basal-like mammary tumor progression; (b) emphasize that alpha(6) integrins function via CD151 linkage in the context of tetraspanin-enriched microdomains; and (c) point to potential relevance of CD151 as a high-priority therapeutic target, with relative selectivity (compared with laminin-binding integrins) for pathologic rather than normal physiology.
Figures





Similar articles
-
Tetraspanin CD151 regulates growth of mammary epithelial cells in three-dimensional extracellular matrix: implication for mammary ductal carcinoma in situ.Cancer Res. 2010 Jun 1;70(11):4698-708. doi: 10.1158/0008-5472.CAN-09-4330. Epub 2010 May 25. Cancer Res. 2010. PMID: 20501858 Free PMC article.
-
Expression of the palmitoylation-deficient CD151 weakens the association of alpha 3 beta 1 integrin with the tetraspanin-enriched microdomains and affects integrin-dependent signaling.J Biol Chem. 2002 Oct 4;277(40):36991-7000. doi: 10.1074/jbc.M205265200. Epub 2002 Jul 10. J Biol Chem. 2002. PMID: 12110679
-
Tetraspanin CD151 regulates transforming growth factor beta signaling: implication in tumor metastasis.Cancer Res. 2010 Jul 15;70(14):6059-70. doi: 10.1158/0008-5472.CAN-09-3497. Epub 2010 Jun 22. Cancer Res. 2010. PMID: 20570898 Free PMC article.
-
CD151 in cancer progression and metastasis: a complex scenario.Lab Invest. 2014 Jan;94(1):41-51. doi: 10.1038/labinvest.2013.136. Epub 2013 Nov 18. Lab Invest. 2014. PMID: 24247563 Review.
-
β1-integrins signaling and mammary tumor progression in transgenic mouse models: implications for human breast cancer.Breast Cancer Res. 2011;13(6):229. doi: 10.1186/bcr2905. Epub 2011 Nov 30. Breast Cancer Res. 2011. PMID: 22264244 Free PMC article. Review.
Cited by
-
Tetraspanin CD151 mediates papillomavirus type 16 endocytosis.J Virol. 2013 Mar;87(6):3435-46. doi: 10.1128/JVI.02906-12. Epub 2013 Jan 9. J Virol. 2013. PMID: 23302890 Free PMC article.
-
CD151 restricts the α6 integrin diffusion mode.J Cell Sci. 2012 Mar 15;125(Pt 6):1478-87. doi: 10.1242/jcs.093963. Epub 2012 Feb 10. J Cell Sci. 2012. PMID: 22328509 Free PMC article.
-
The histone H2B-specific ubiquitin ligase RNF20/hBRE1 acts as a putative tumor suppressor through selective regulation of gene expression.Genes Dev. 2008 Oct 1;22(19):2664-76. doi: 10.1101/gad.1703008. Genes Dev. 2008. PMID: 18832071 Free PMC article.
-
CD151 promotes cell metastasis via activating TGF-β1/Smad signaling in renal cell carcinoma.Oncotarget. 2018 Jan 8;9(17):13313-13323. doi: 10.18632/oncotarget.24028. eCollection 2018 Mar 2. Oncotarget. 2018. PMID: 29568359 Free PMC article.
-
Deletion of Cd151 reduces mammary tumorigenesis in the MMTV/PyMT mouse model.BMC Cancer. 2014 Jul 11;14:509. doi: 10.1186/1471-2407-14-509. BMC Cancer. 2014. PMID: 25012362 Free PMC article.
References
-
- Hemler ME. Tetraspanin proteins mediate cellular penetration, invasion and fusion events, and define a novel type of membrane microdomain. Ann Rev Cell Dev Biol. 2003;19:397–422. - PubMed
-
- Sincock PM, Fitter S, Parton RG, Berndt MC, Gamble JR, Ashman LK. PETA-3/CD151, a member of the transmembrane 4 superfamily, is localised to the plasma membrane and endocytic system of endothelial cells, associates with multiple integrins and modulates cell function. J Cell Sci. 1999;112:833–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
