

Specificity of amyloid precursor-like protein 2 interactions with MHC class I molecules
- PMID: 18452037
- PMCID: PMC2683759
- DOI: 10.1007/s00251-008-0296-0
Specificity of amyloid precursor-like protein 2 interactions with MHC class I molecules
Abstract
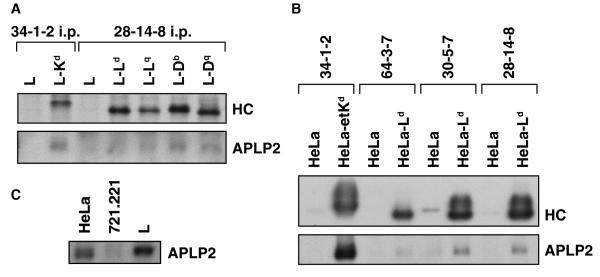
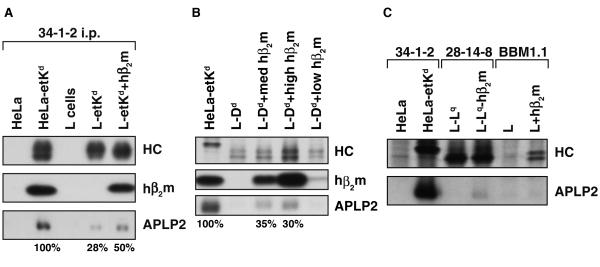
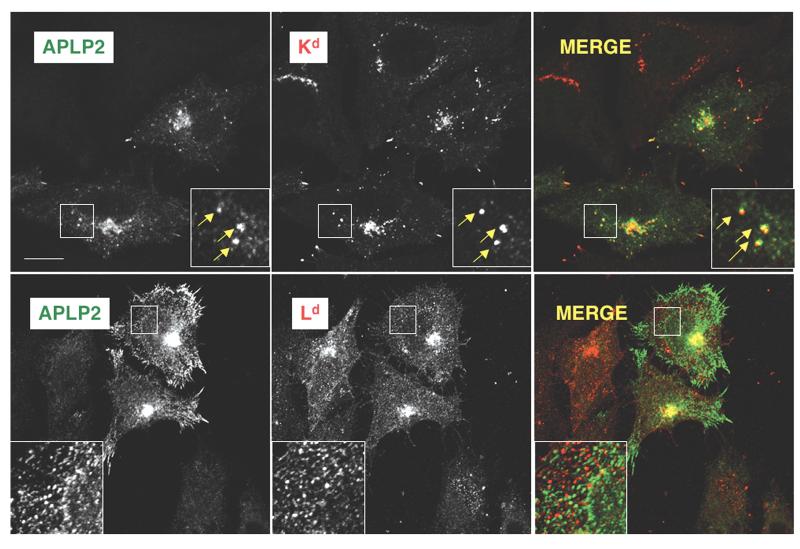
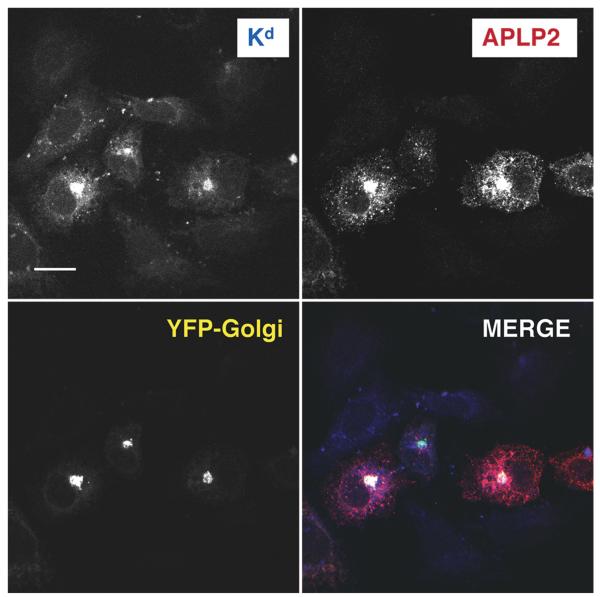
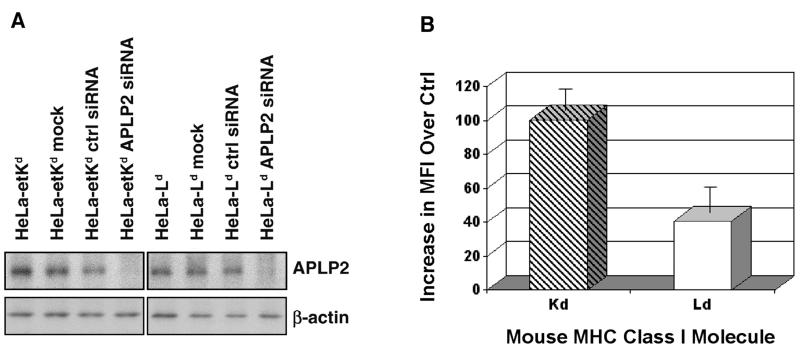
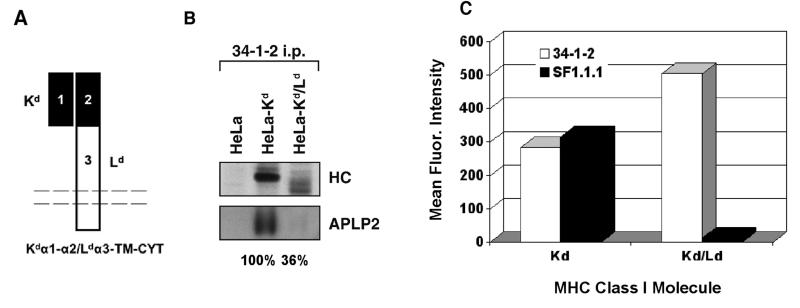
The ubiquitously expressed amyloid precursor-like protein 2 (APLP2) has been previously found to regulate cell surface expression of the major histocompatibility complex (MHC) class I molecule K(d) and bind strongly to K(d). In the study reported here, we demonstrated that APLP2 binds, in varied degrees, to several other mouse MHC class I allotypes and that the ability of APLP2 to affect cell surface expression of an MHC class I molecule is not limited to K(d). L(d), like K(d), was found associated with APLP2 in the Golgi, but K(d) was also associated with APLP2 within intracellular vesicular structures. We also investigated the effect of beta(2)m on APLP2/MHC interaction and found that human beta(2)m transfection increased the association of APLP2 with mouse MHC class I molecules, likely by affecting H2 class I heavy chain conformation. APLP2 was demonstrated to bind specifically to the conformation of L(d) having folded outer domains, consistent with our previous results with K(d) and indicating APLP2 interacts with the alpha1alpha2 region on each of these H2 class I molecules. Furthermore, we observed that binding to APLP2 involved the MHC alpha3/transmembrane/cytoplasmic region, suggesting that conserved as well as polymorphic regions of the H2 class I molecule may participate in interaction with APLP2. In summary, we demonstrated that APLP2's binding, co-localization pattern, and functional impact vary among H2 class I molecules and that APLP2/MHC association is influenced by multiple domains of the MHC class I heavy chain and by beta(2)m's effects on the conformation of the heavy chain.
Figures
References
-
- Achour A, Michaelsson J, Harris RA, Ljunggren H-G, Karre K, Schnieder G, Scandalova T. Structural basis of the differential stability and receptor-specificity of H-2Db in complex with murine versus human β2-microglobulin. J Mol Biol. 2006;356:382–396. - PubMed
-
- Benoit LA, Tan R. Xenogeneic β2-microglobulin substitution affects functional binding of MHC class I molecules by CD8+ T cells. J Immunol. 2007;179:3588–3595. - PubMed
-
- Brodsky FM, Bodmer WF, Parham P. Characterization of a monoclonal anti-beta 2-microglobulin antibody and its use in the genetic and biochemical analysis of major histocompatibility antigens. Eur J Immunol. 1979;9:536–545. - PubMed
-
- Cappai R, Mok SS, Galatis D, Tucker DF, Henry A, Beyreuther K, Small DH, Masters CL. Recombinant human amyloid precursor-like protein 2 (APLP2) expressed in the yeast Pichia pastoris can stimulate neurite outgrowth. FEBS Lett. 1999;442:95–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
