

Carrageenan-induced NFkappaB activation depends on distinct pathways mediated by reactive oxygen species and Hsp27 or by Bcl10
- PMID: 18452717
- PMCID: PMC3698247
- DOI: 10.1016/j.bbagen.2008.03.019
Carrageenan-induced NFkappaB activation depends on distinct pathways mediated by reactive oxygen species and Hsp27 or by Bcl10
Abstract
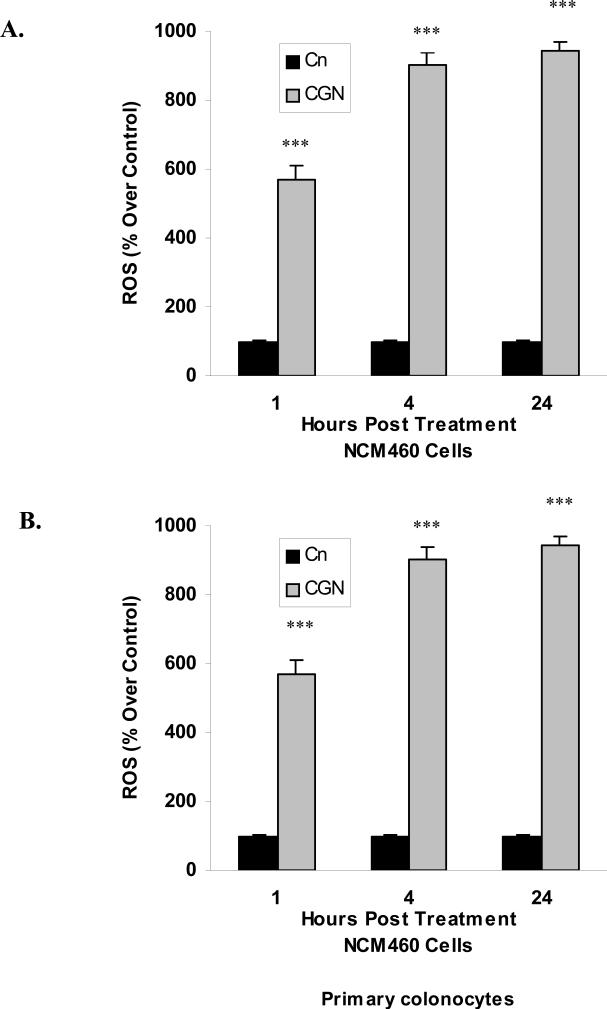
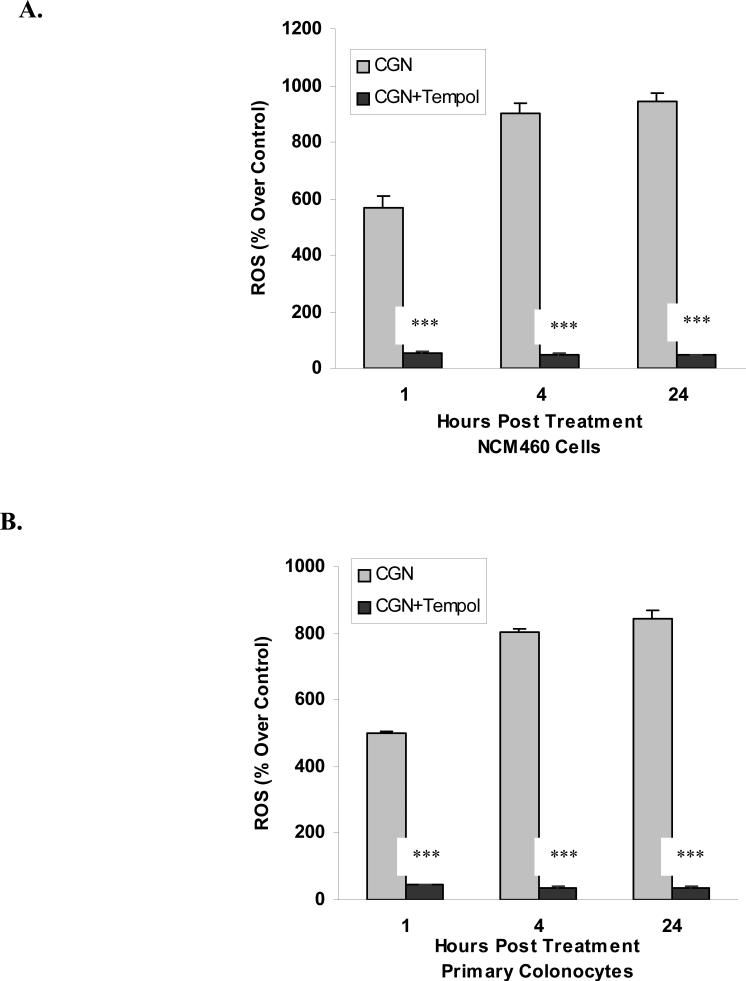
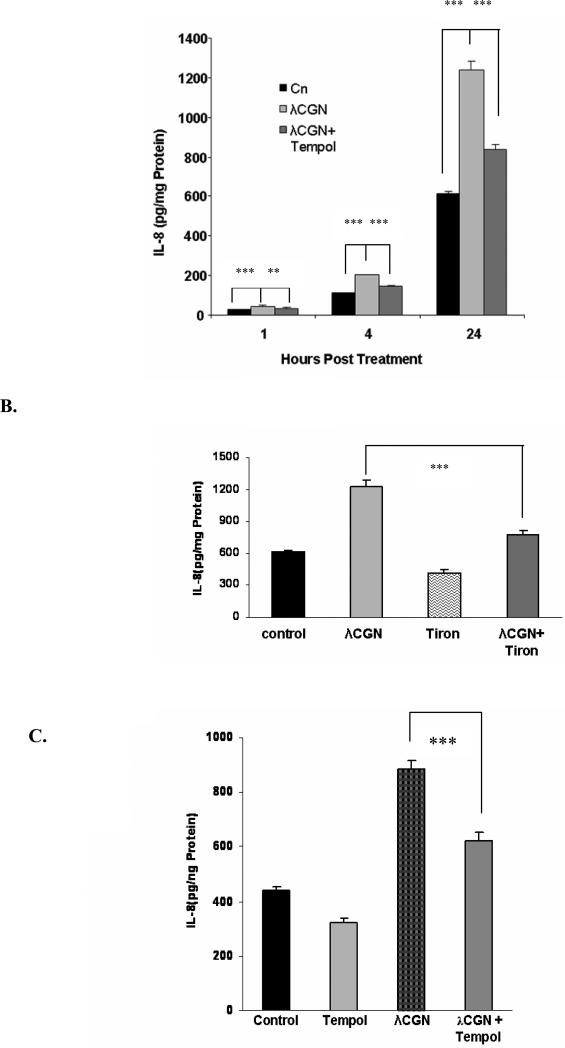
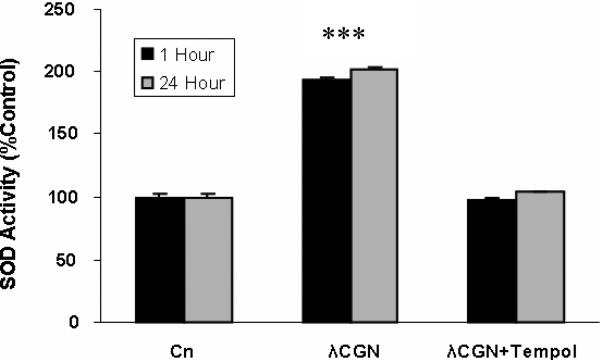
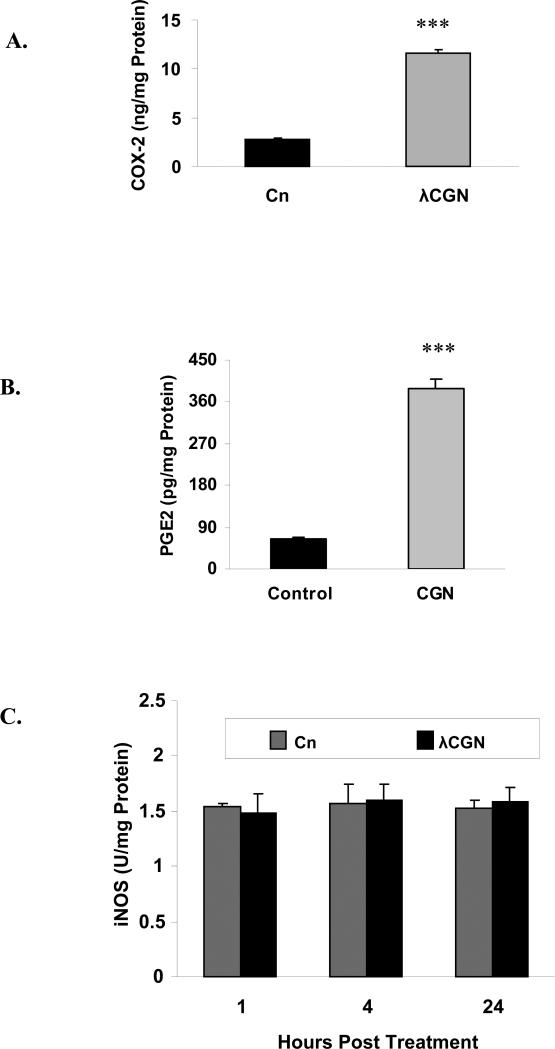
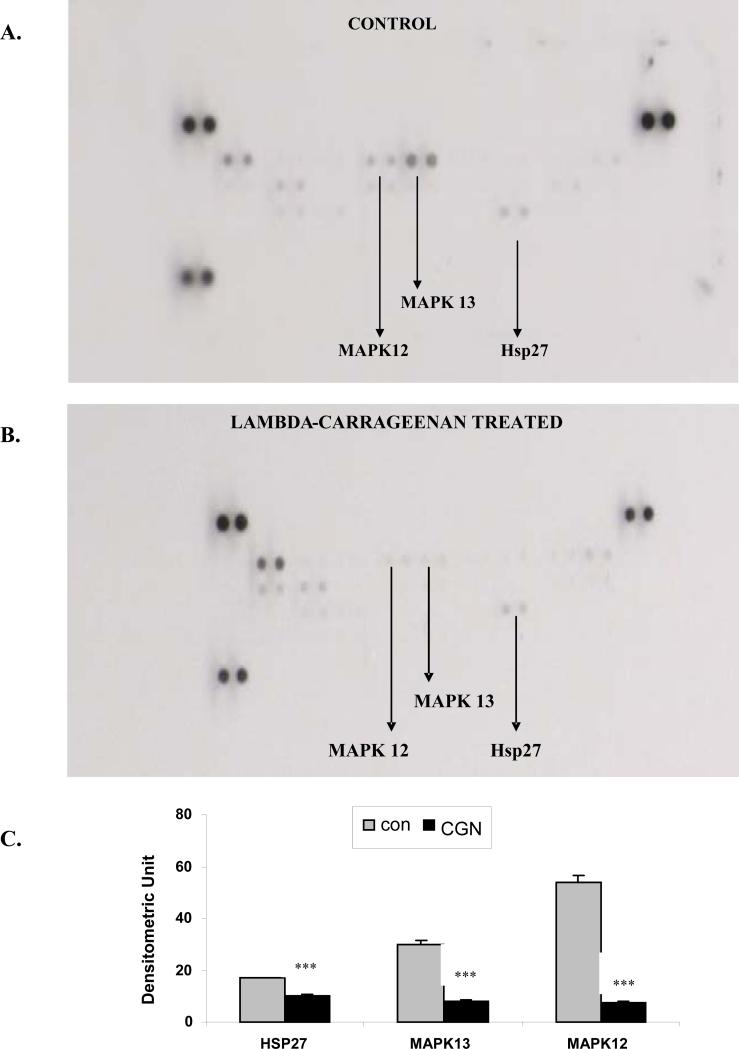
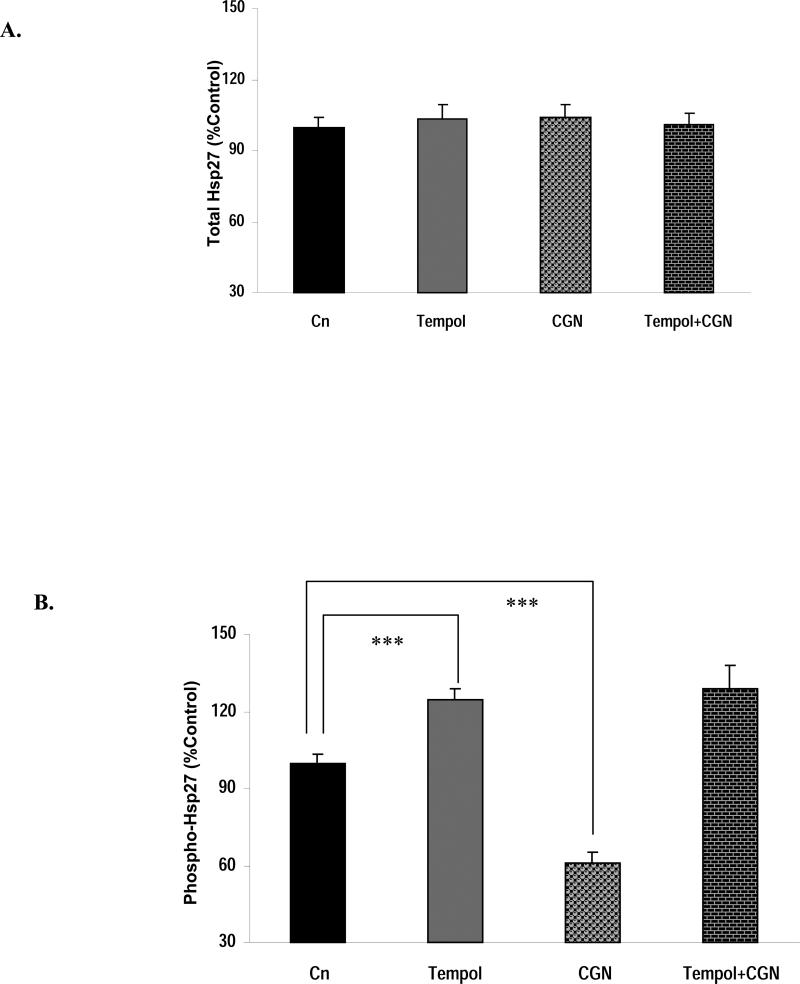
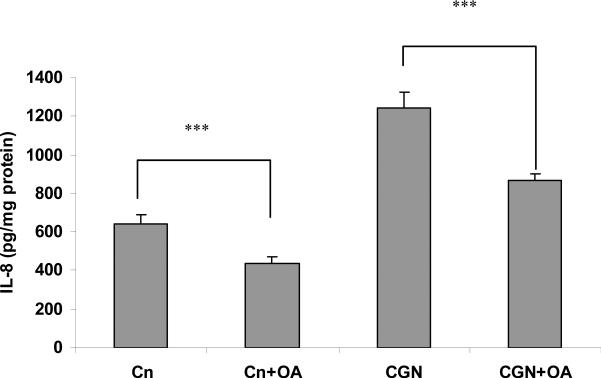
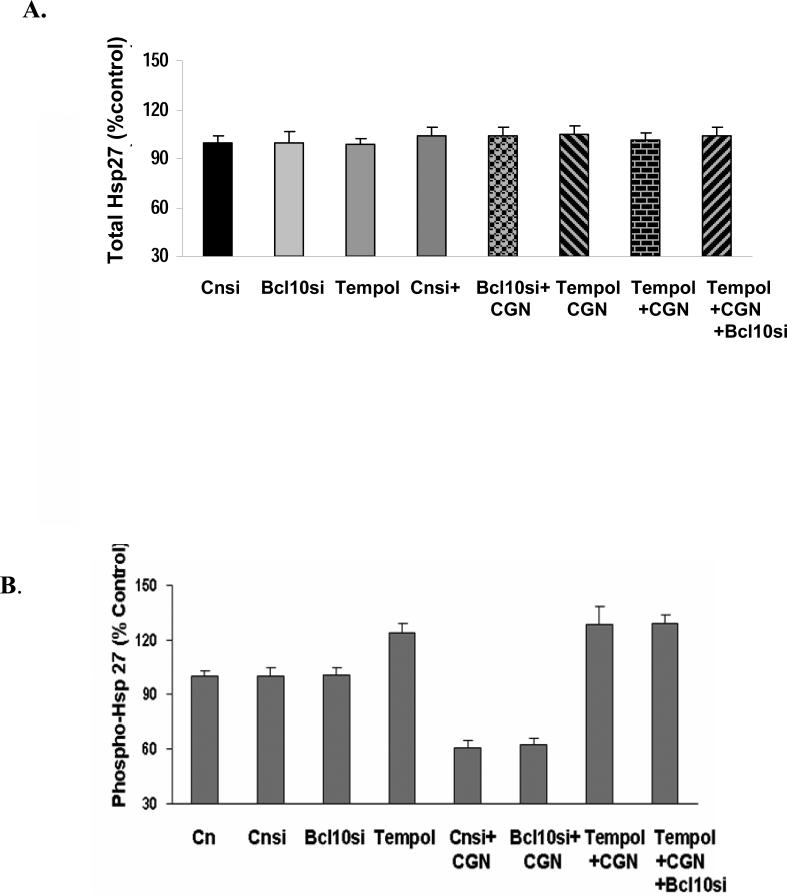
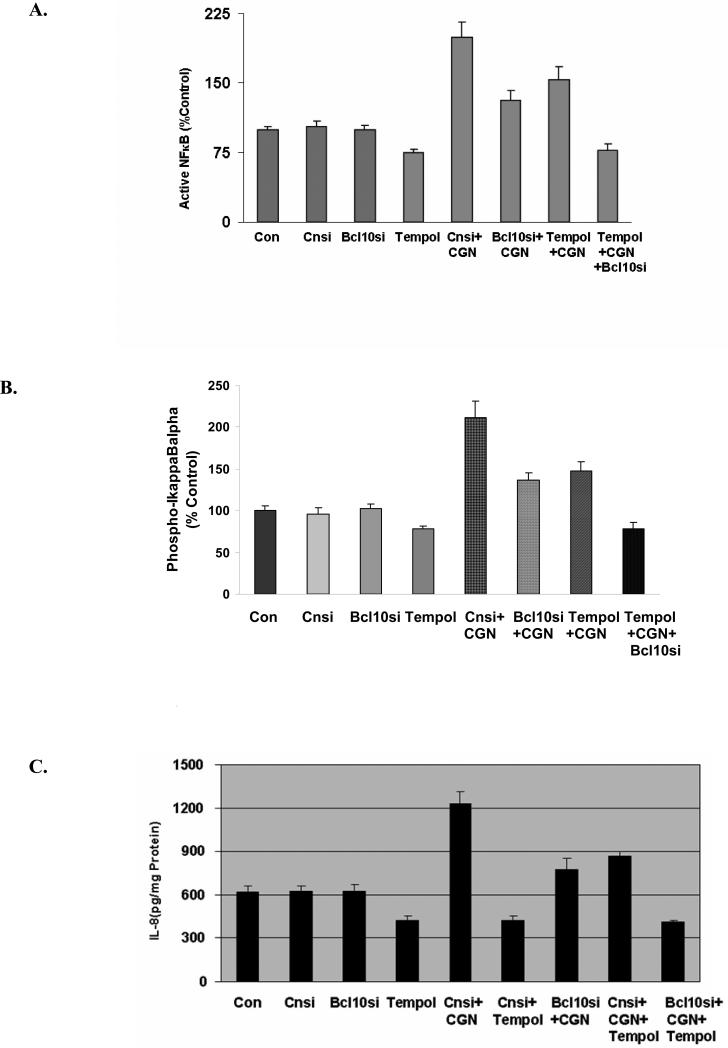
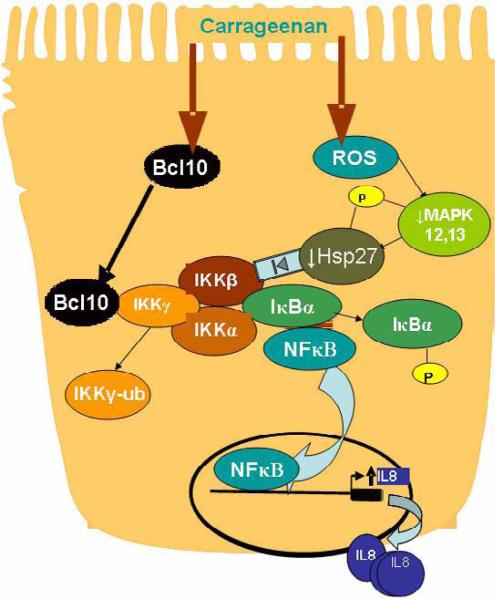
Carrageenans are highly sulfated polysaccharides that are widely used as food additives due to their ability to improve food texture. They are also widely recognized for their ability to induce inflammation in animal models of colitis. Recently, we reported that carrageenan (CGN) activated a pathway of innate immunity in human colonic epithelial cells mediated by Bcl10 (B-cell CLL/lymphoma 10). However, increases in phospho-IkappaBalpha and Interleukin-8 (IL-8) were not completely inhibited by silencing Bcl10, suggesting that CGN also influenced another mechanism, or mechanisms, of inflammation. In this report, we demonstrate that CGN increases production of reactive oxygen species (ROS) in human colonic epithelial cells. The combination of ROS quenching by the free radical scavenger Tempol and of Bcl10 silencing by siRNA completely inhibited the CGN-induced increases in nuclear NFkappaB (p65), phospho-IkappaBalpha, and secretion of IL-8. The CGN-induced increase in ROS was associated with declines in phosphorylation of MAPK 12 (p38gamma), MAPK 13 (p38delta), and heat-shock protein (Hsp) 27. The CGN-induced decline in phospho-Hsp27 was reversed by co-administration of Tempol (100 nM), but unaffected by silencing Bcl10. Since Hsp27 phosphorylation is inversely associated with phosphorylation of the IkappaBalpha kinase (IKK) signalosome, CGN exposure appears to affect the IKK signalosome by both the catalytic component, mediated by ROS-phospho-Hsp27, and the regulatory component, mediated by Bcl10 interaction with IKKgamma (Nemo). Hence, the CGN-activated inflammatory cascades related to innate immunity and to generation of ROS may be integrated at the level of the IKK signalosome.
Figures
Similar articles
-
B-cell CLL/lymphoma 10 (BCL10) is required for NF-kappaB production by both canonical and noncanonical pathways and for NF-kappaB-inducing kinase (NIK) phosphorylation.J Biol Chem. 2010 Jan 1;285(1):522-30. doi: 10.1074/jbc.M109.050815. Epub 2009 Nov 6. J Biol Chem. 2010. PMID: 19897484 Free PMC article.
-
Lipopolysaccharide activates NF-kappaB by TLR4-Bcl10-dependent and independent pathways in colonic epithelial cells.Am J Physiol Gastrointest Liver Physiol. 2008 Oct;295(4):G784-90. doi: 10.1152/ajpgi.90434.2008. Epub 2008 Aug 21. Am J Physiol Gastrointest Liver Physiol. 2008. PMID: 18718996
-
ROS, Hsp27, and IKKbeta mediate dextran sodium sulfate (DSS) activation of IkappaBa, NFkappaB, and IL-8.Inflamm Bowel Dis. 2009 May;15(5):673-83. doi: 10.1002/ibd.20821. Inflamm Bowel Dis. 2009. PMID: 19085995 Free PMC article.
-
The effect of reactive oxygen species on the synthesis of prostanoids from arachidonic acid.J Physiol Pharmacol. 2013 Aug;64(4):409-21. J Physiol Pharmacol. 2013. PMID: 24101387 Review.
-
The Role of Carrageenan in Inflammatory Bowel Diseases and Allergic Reactions: Where Do We Stand?Nutrients. 2021 Sep 27;13(10):3402. doi: 10.3390/nu13103402. Nutrients. 2021. PMID: 34684400 Free PMC article. Review.
Cited by
-
Hyperglycemia and advanced glycosylation end products suppress adipocyte apoE expression: implications for adipocyte triglyceride metabolism.Am J Physiol Endocrinol Metab. 2010 Oct;299(4):E615-23. doi: 10.1152/ajpendo.00273.2010. Epub 2010 Jul 20. Am J Physiol Endocrinol Metab. 2010. PMID: 20647555 Free PMC article.
-
Increased expression of colonic Wnt9A through Sp1-mediated transcriptional effects involving arylsulfatase B, chondroitin 4-sulfate, and galectin-3.J Biol Chem. 2014 Jun 20;289(25):17564-75. doi: 10.1074/jbc.M114.561589. Epub 2014 Apr 28. J Biol Chem. 2014. PMID: 24778176 Free PMC article.
-
Carrageenan as a Potential Factor of Inflammatory Bowel Diseases.Nutrients. 2024 Apr 30;16(9):1367. doi: 10.3390/nu16091367. Nutrients. 2024. PMID: 38732613 Free PMC article. Review.
-
Animal models of colitis-associated carcinogenesis.J Biomed Biotechnol. 2011;2011:342637. doi: 10.1155/2011/342637. Epub 2011 Jan 12. J Biomed Biotechnol. 2011. PMID: 21274454 Free PMC article. Review.
-
Effects of tempol and redox-cycling nitroxides in models of oxidative stress.Pharmacol Ther. 2010 May;126(2):119-45. doi: 10.1016/j.pharmthera.2010.01.003. Epub 2010 Feb 11. Pharmacol Ther. 2010. PMID: 20153367 Free PMC article. Review.
References
-
- Galili U. The alpha-gal epitope and the anti-Gal antibody in xenotransplantation and in cancer immunotherapy. Immunol. Cell Biol. 2005;83:674–86. - PubMed
-
- Sandrin MS, Osman N, McKenzie IFC. Transgenic approaches for the reduction in expression of GALα(1,3)GAL for xenotransplantation. Front. Biosci. 1997;2:e1–11. - PubMed
-
- Baumann BC, Stussi G, Huggel K, Rieben R, Seebach JD. Reactivity of human natural antibodies to endothelial cells from Galpha(1,3)Gal-deficient pigs. Transplantation. 2007;83:193–201. - PubMed
-
- Borthakur A, Bhattacharyya S, Dudeja PK, Tobacman JK. Carrageenan induces NFκB activation and interleukin-8 production in normal human colonic epithelial cells. Am. J. Physiol.: Gastrointest. Liver Physiol. 2007;292:G829–G838. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
