

Characterization of Dnmt3b:thymine-DNA glycosylase interaction and stimulation of thymine glycosylase-mediated repair by DNA methyltransferase(s) and RNA
- PMID: 18452947
- PMCID: PMC2705441
- DOI: 10.1016/j.jmb.2008.02.049
Characterization of Dnmt3b:thymine-DNA glycosylase interaction and stimulation of thymine glycosylase-mediated repair by DNA methyltransferase(s) and RNA
Abstract
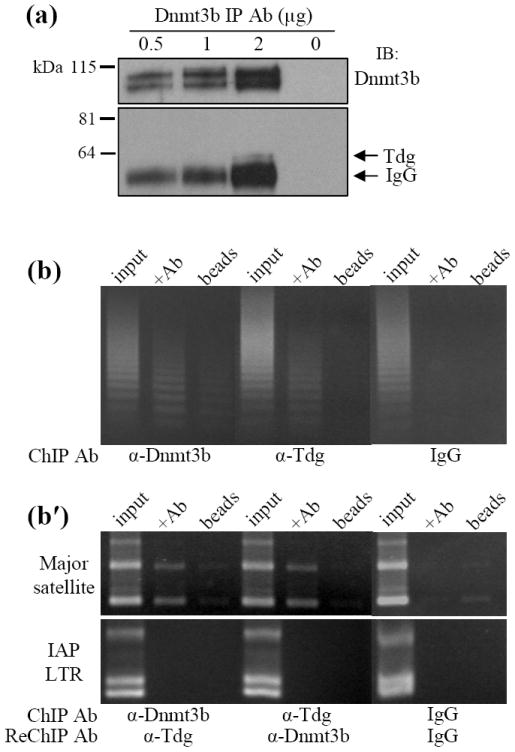
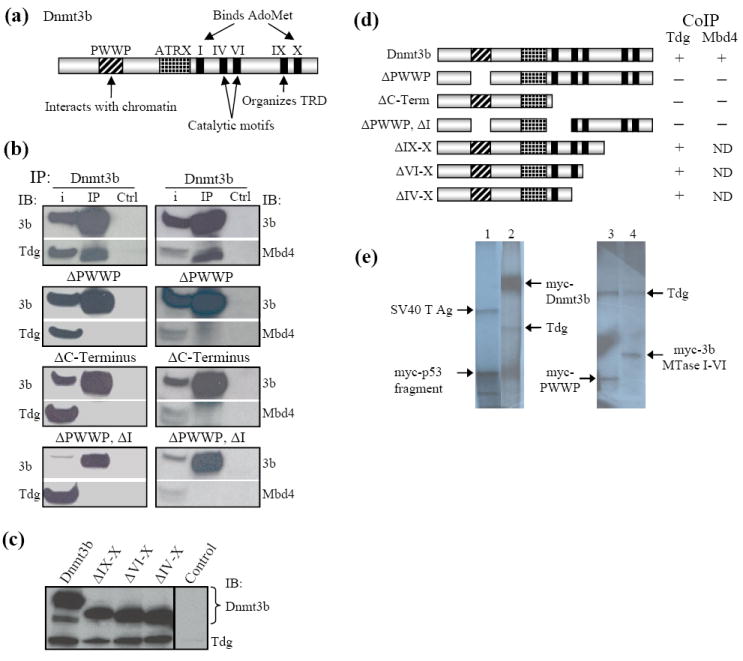
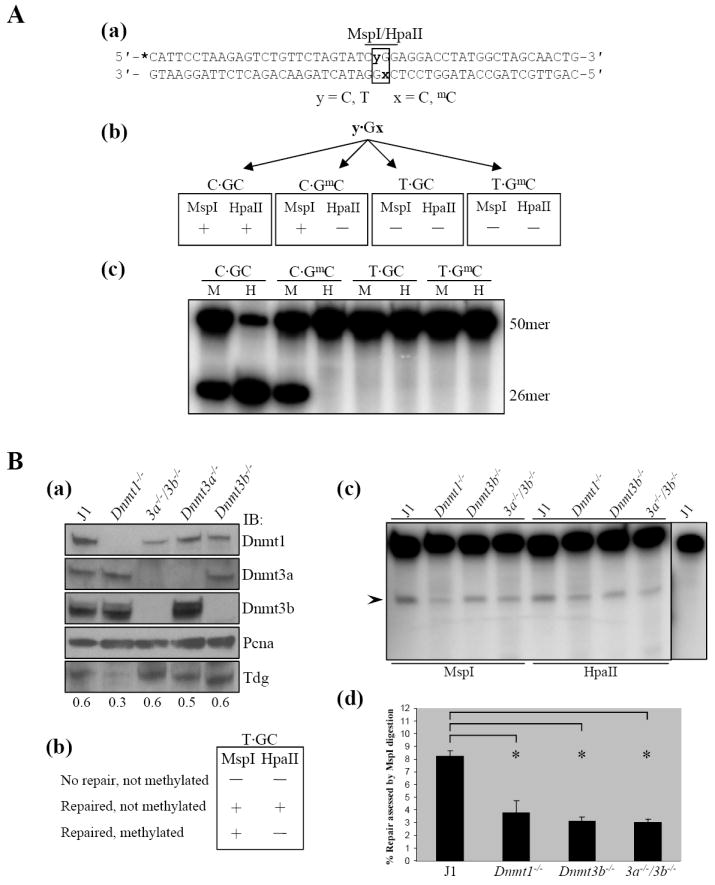
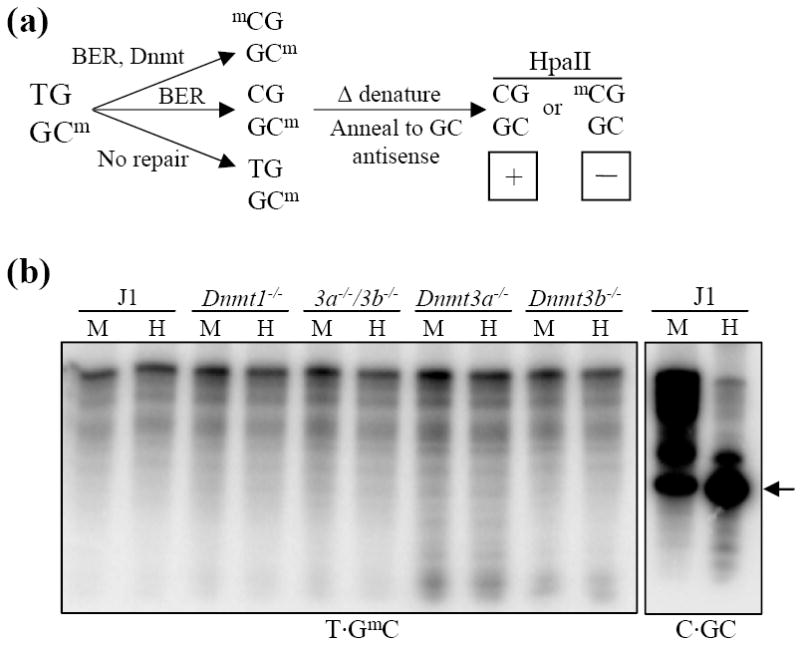
Methylation of cytosine residues in CpG dinucleotides plays an important role in epigenetic regulation of gene expression and chromatin structure/stability in higher eukaryotes. DNA methylation patterns are established and maintained at CpG dinucleotides by DNA methyltransferases (Dnmt1, Dnmt3a, and Dnmt3b). In mammals and many other eukaryotes, the CpG dinucleotide is underrepresented in the genome. This loss is postulated to be the result of unrepaired deamination of cytosine and 5-methylcytosine to uracil and thymine, respectively. Two thymine glycosylases are believed to reduce the impact of 5-methylcytosine deamination. G/T mismatch-specific thymine-DNA glycosylase (Tdg) and methyl-CpG binding domain protein 4 can both excise uracil or thymine at U.G and T.G mismatches to initiate base excision repair. Here, we report the characterization of interactions between Dnmt3b and both Tdg and methyl-CpG binding domain protein 4. Our results demonstrate (1) that both Tdg and Dnmt3b are colocalized to heterochromatin and (2) reduction of T.G mismatch repair efficiency upon loss of DNA methyltransferase expression, as well as a requirement for an RNA component for correct T.G mismatch repair.
Figures

References
-
- Chen RZ, Pettersson U, Beard C, Jackson-Grusby L, Jaenisch R. DNA hypomethylation leads to elevated mutation rates. Nature. 1998;395:89–93. - PubMed
-
- Rizwana R, Hahn PJ. CpG methylation reduces genomic instability. J Cell Sci. 1999;112(Pt 24):4513–4519. - PubMed
-
- Li E. Chromatin modification and epigenetic reprogramming in mammalian development. Nat Rev Genet. 2002;3:662–673. - PubMed
-
- Okano M, Bell DW, Haber DA, Li E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell. 1999;99:247–257. - PubMed
-
- Okano M, Xie S, Li E. Cloning and characterization of a family of novel mammalian DNA (cytosine-5) methyltransferases. Nat Genet. 1998;19:219–220. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
