

Shotgun lipidomics reveals the temporally dependent, highly diversified cardiolipin profile in the mammalian brain: temporally coordinated postnatal diversification of cardiolipin molecular species with neuronal remodeling
- PMID: 18454555
- PMCID: PMC2507872
- DOI: 10.1021/bi7023282
Shotgun lipidomics reveals the temporally dependent, highly diversified cardiolipin profile in the mammalian brain: temporally coordinated postnatal diversification of cardiolipin molecular species with neuronal remodeling
Abstract
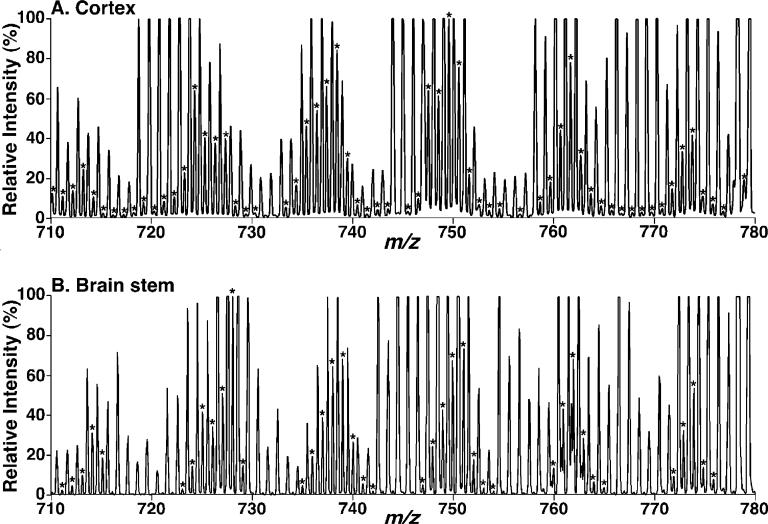
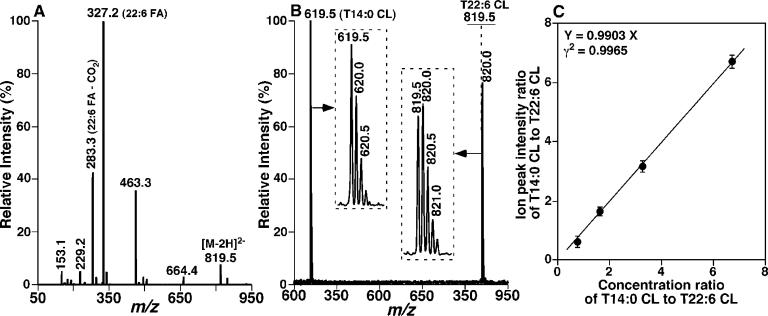
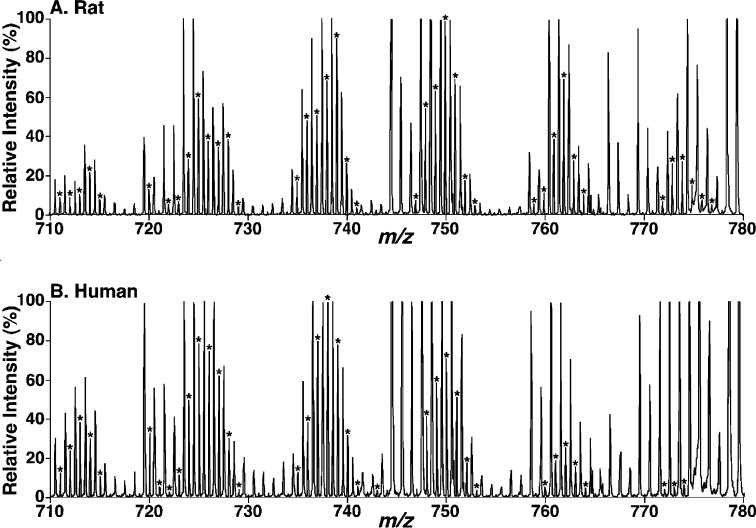
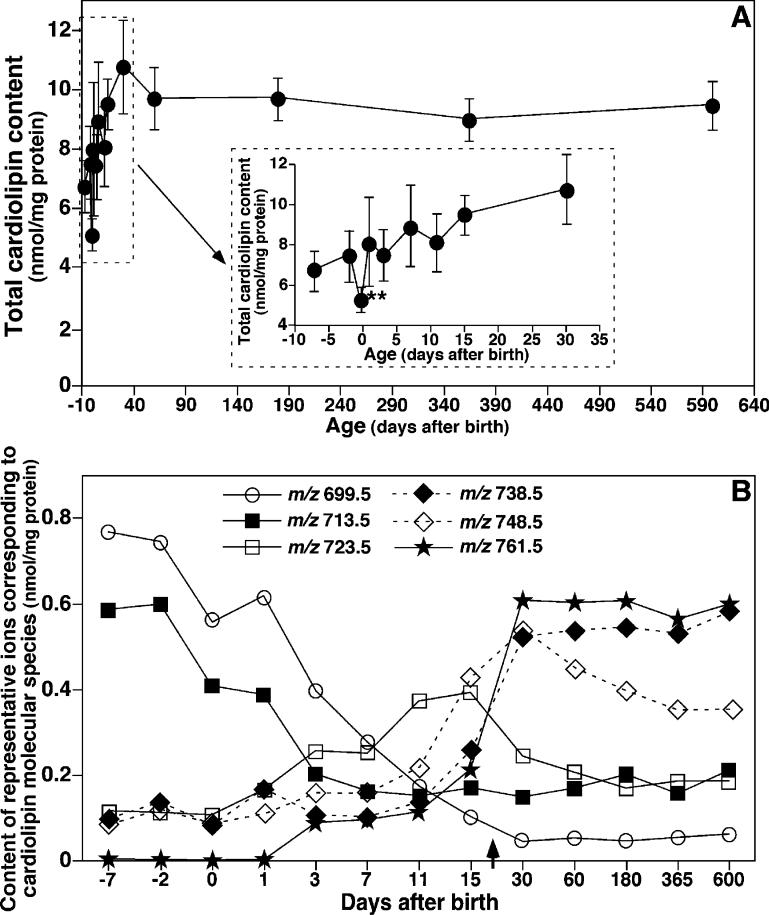
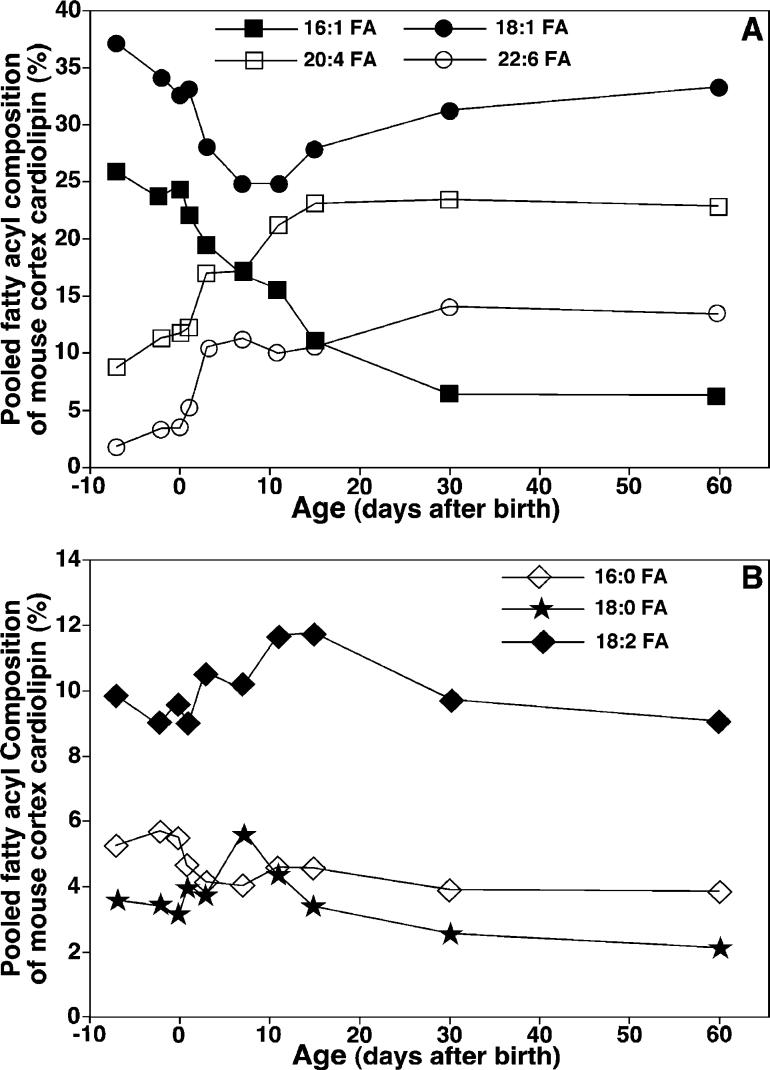
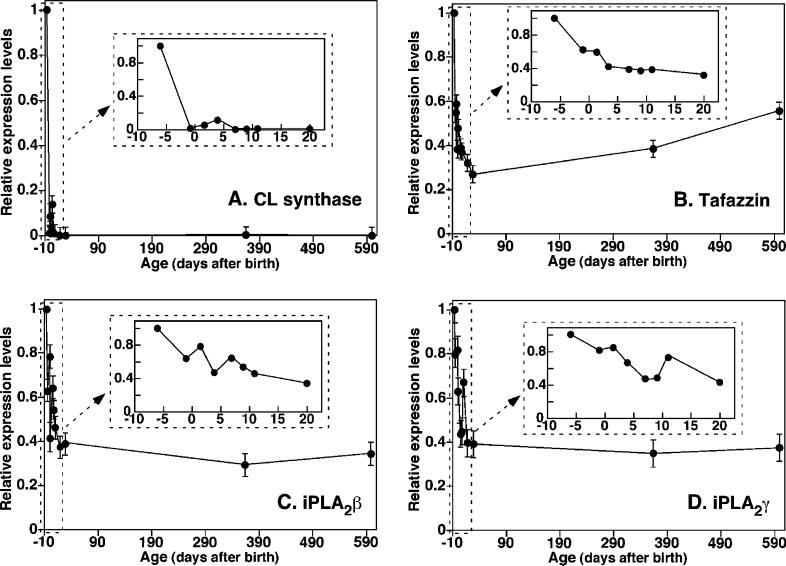
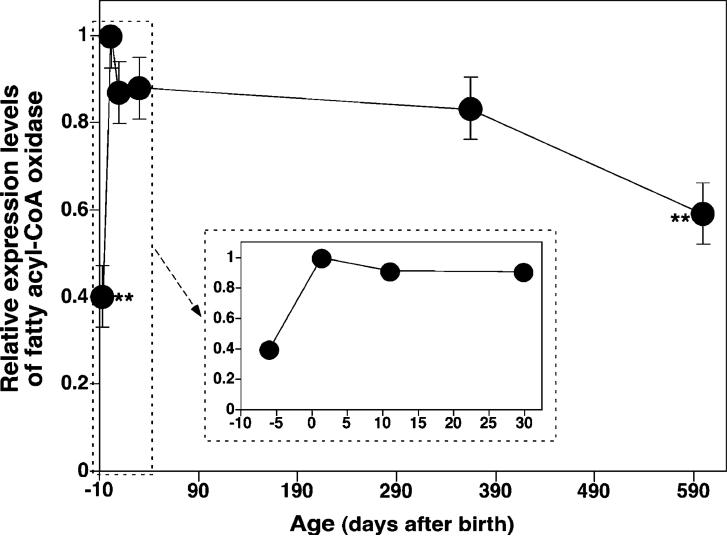
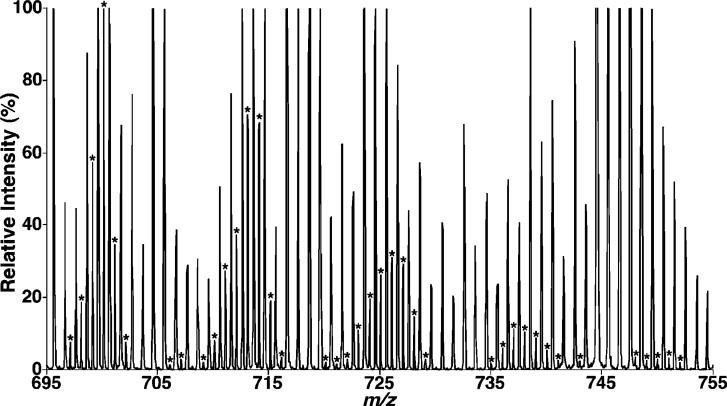
Large-scale neuronal remodeling through apoptosis occurs shortly after birth in all known mammalian species. Apoptosis, in large part, depends upon critical interactions between mitochondrial membranes and cytochrome c. Herein, we examined the hypothesis that the large-scale reorganization of neuronal circuitry after birth is accompanied by profound alterations in cardiolipin (CL) content and molecular species distribution. During embryonic development, over 100 CL molecular species were identified and quantitated in murine neuronal tissues. The embryonic CL profile was notable for the presence of abundant amounts of relatively short aliphatic chains (e.g., palmitoleic and oleic acids). In sharp contrast, after birth, the CL profile contained a remarkably complex repertoire of CL molecular species, in which the signaling fatty acids (i.e., arachidonic and docosahexaenoic acids) were markedly increased. These results identify the rapid remodeling of CL in the perinatal period with resultant alterations in the physical properties of the mitochondrial membrane. The complex distribution of aliphatic chains in the neuronal CL pool is separate and distinct from that in other organs (e.g., heart, liver, etc.), where CL molecular species contain predominantly only one major type of aliphatic chain (e.g., linoleic acid). Analyses of mRNA levels by real-time quantitative polymerase chain reactions suggested that the alterations in CL content were due to the combined effects of both attenuation of de novo CL biosynthesis and decreased remodeling of CL. Collectively, these results provide a new perspective on the complexity of CL in neuronal signaling, mitochondrial bioenergetics, and apoptosis.
Figures
References
-
- Schlame M, Rua D, Greenberg ML. The biosynthesis and functional role of cardiolipin. Prog. Lipid Res. 2000;39:257–288. - PubMed
-
- Hoch FL. Cardiolipins and biomembrane function. Biochim. Biophys. Acta. 1992;1113:71–133. - PubMed
-
- Hatch GM. Cell biology of cardiac mitochondrial phospholipids. Biochem. Cell Biol. 2004;82:99–112. - PubMed
-
- Petrosillo G, Ruggiero FM, Paradies G. Role of reactive oxygen species and cardiolipin in the release of cytochrome c from mitochondria. FASEB J. 2003;17:2202–2208. - PubMed
-
- Iverson SL, Orrenius S. The cardiolipin—cytochrome c interaction and the mitochondrial regulation of apoptosis. Arch. Biochem. Biophys. 2004;423:37–46. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
