

Mutations outside the anisomycin-binding site can make ribosomes drug-resistant
- PMID: 18455733
- PMCID: PMC2442718
- DOI: 10.1016/j.jmb.2008.03.075
Mutations outside the anisomycin-binding site can make ribosomes drug-resistant
Abstract
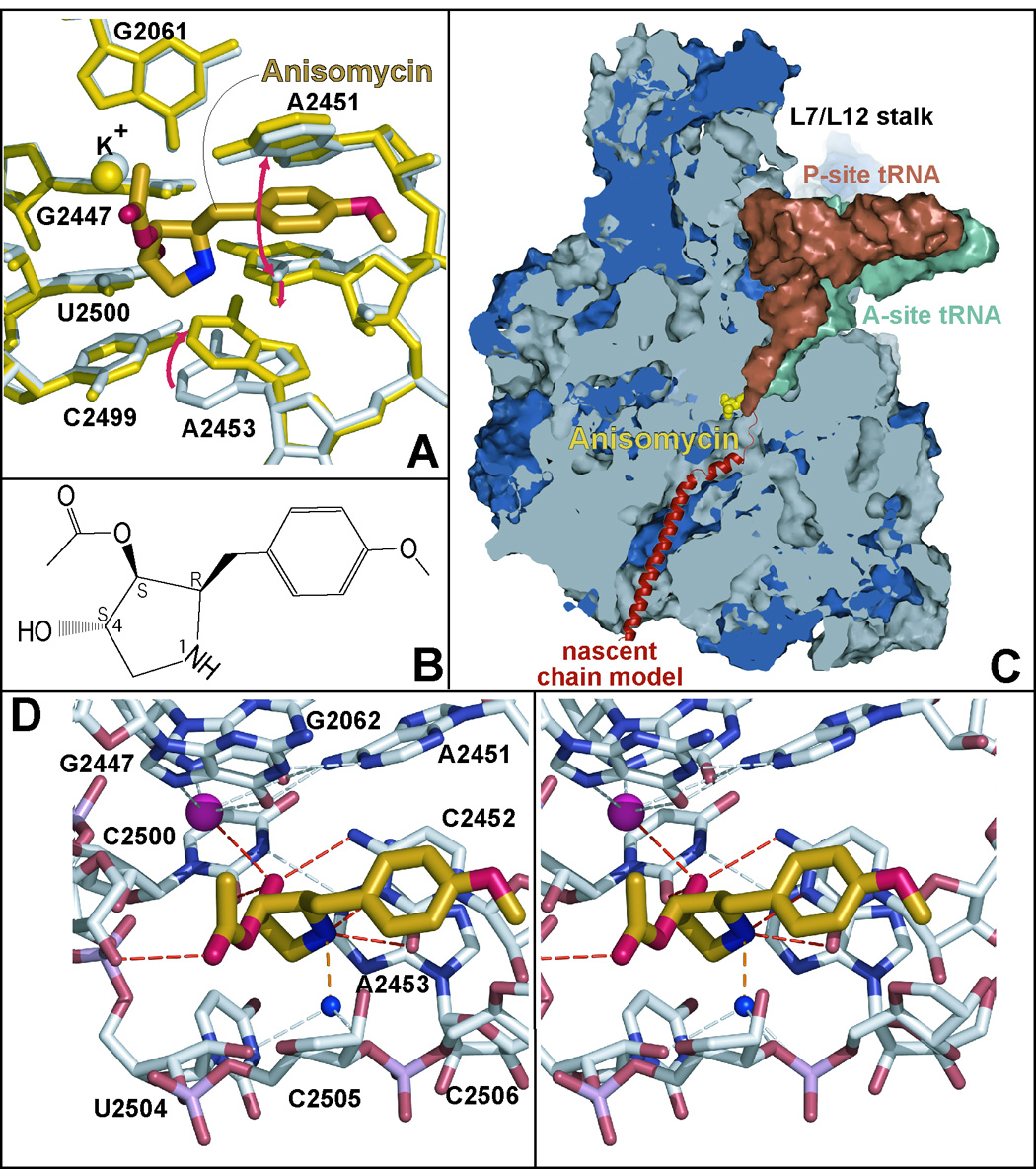
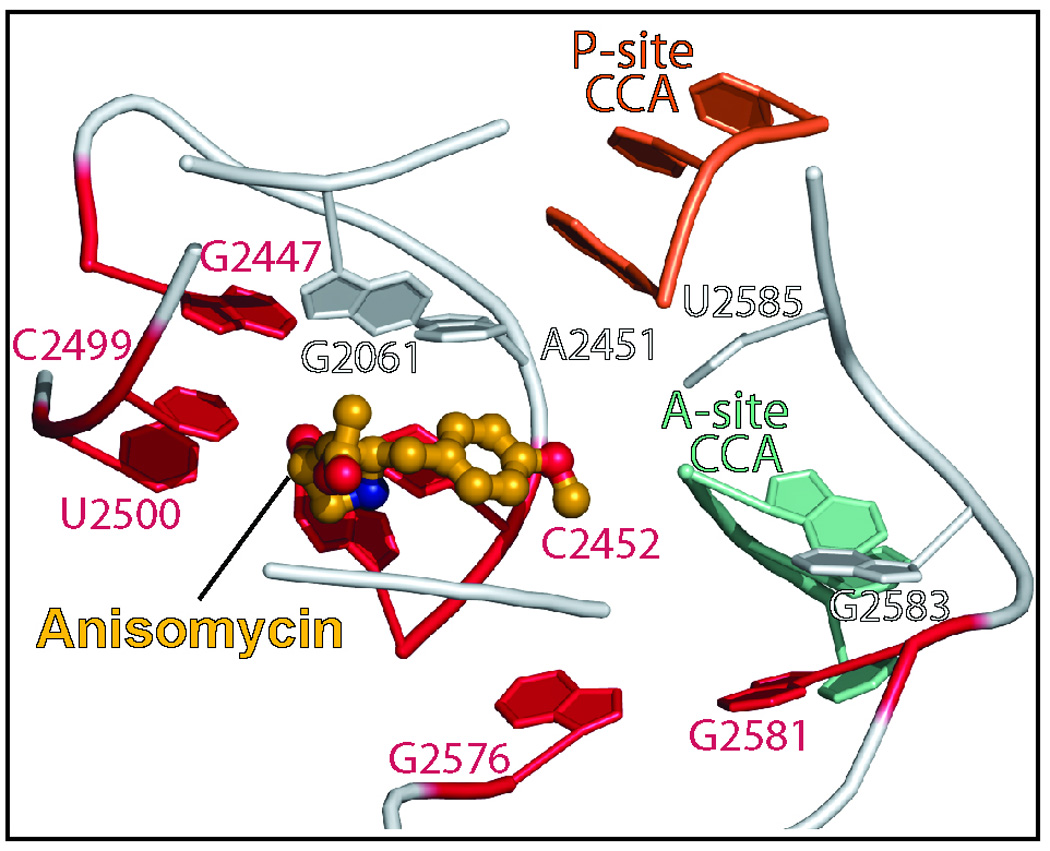
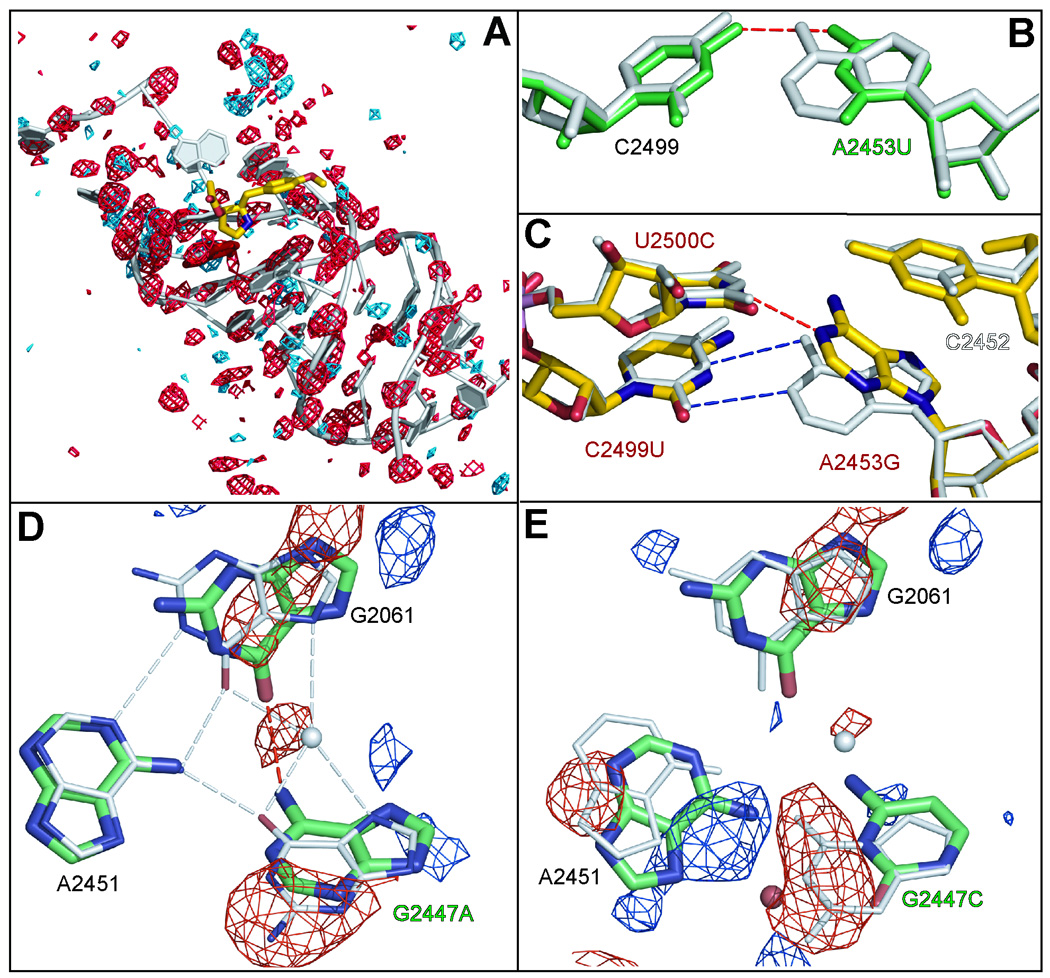
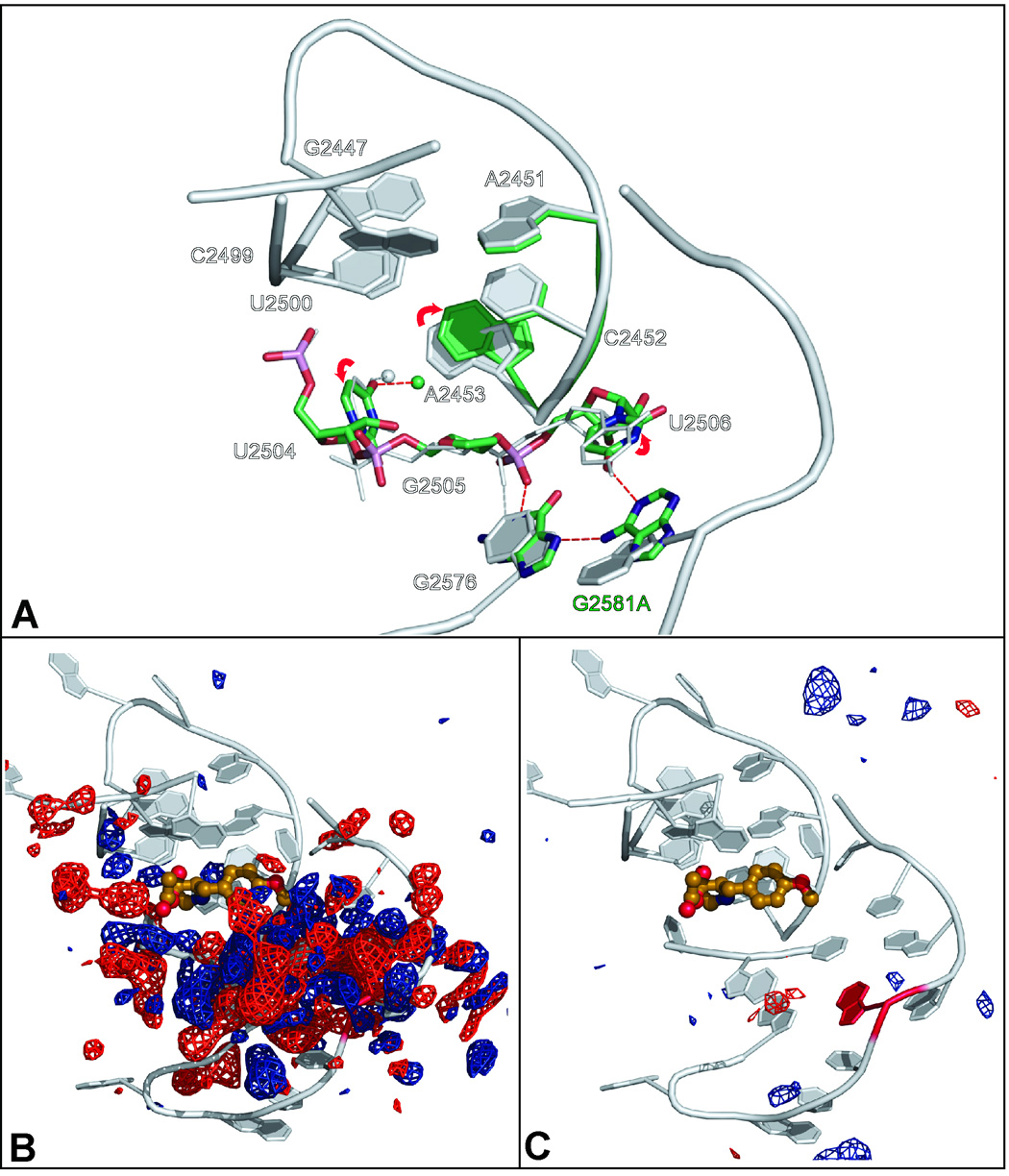
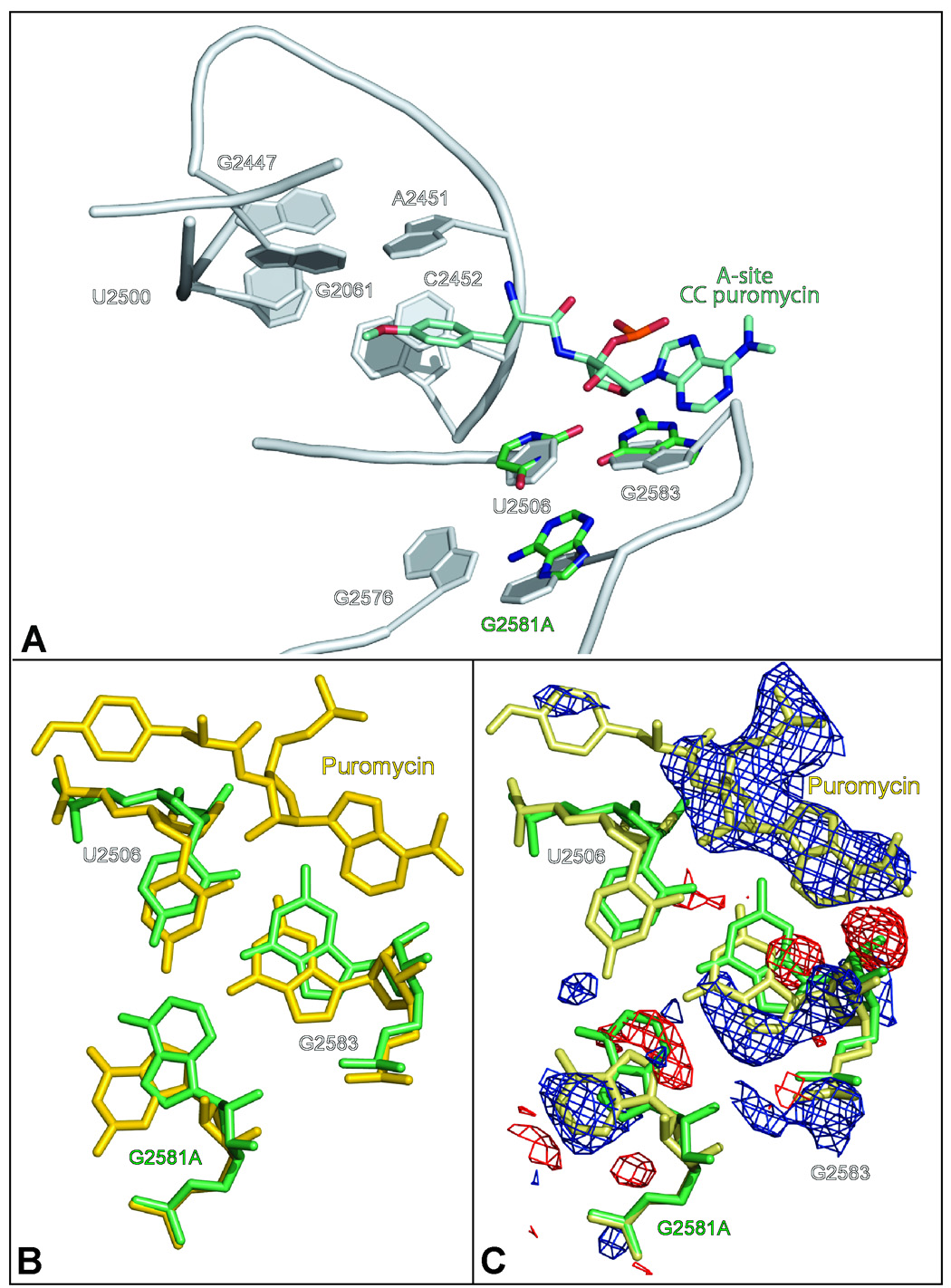
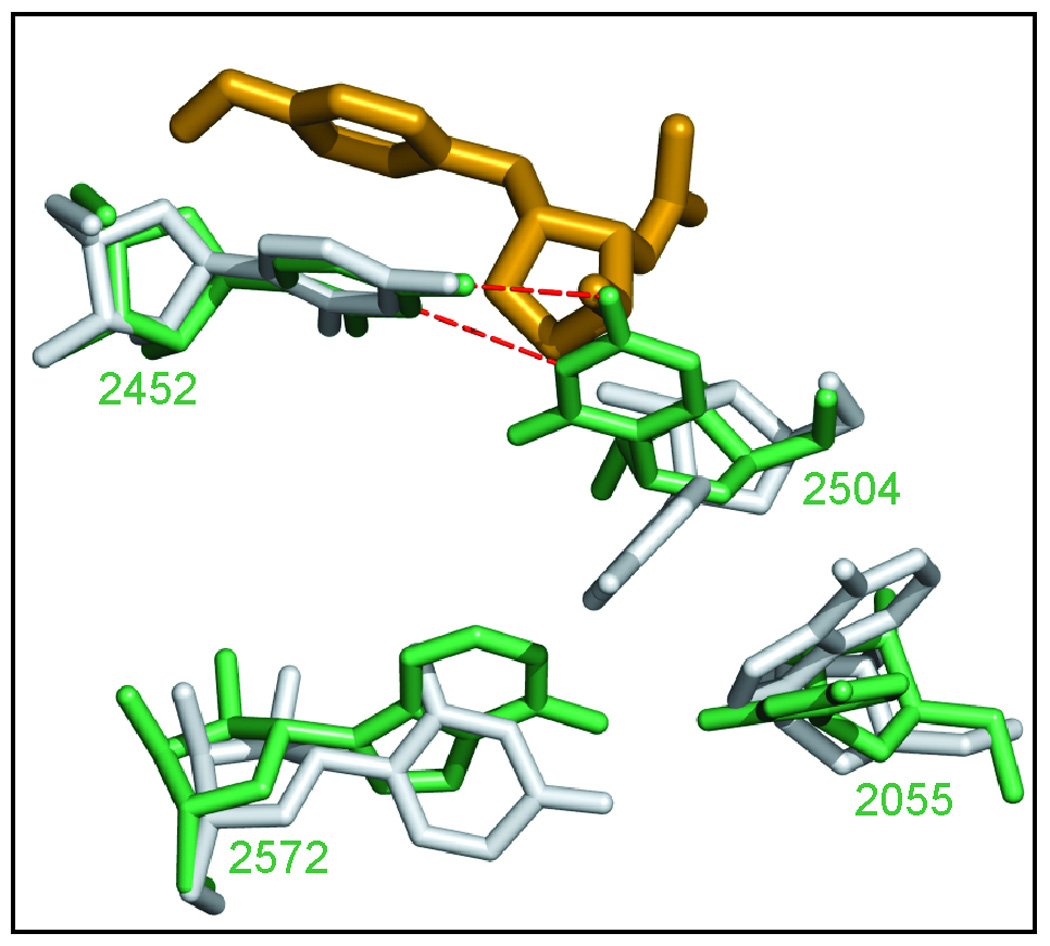
Eleven mutations that make Haloarcula marismortui resistant to anisomycin, an antibiotic that competes with the amino acid side chains of aminoacyl tRNAs for binding to the A-site cleft of the large ribosomal unit, have been identified in 23S rRNA. The correlation observed between the sensitivity of H. marismortui to anisomycin and the affinity of its large ribosomal subunits for the drug indicates that its response to anisomycin is determined primarily by the binding of the drug to its large ribosomal subunit. The structures of large ribosomal subunits containing resistance mutations show that these mutations can be divided into two classes: (1) those that interfere with specific drug-ribosome interactions and (2) those that stabilize the apo conformation of the A-site cleft of the ribosome relative to its drug-bound conformation. The conformational effects of some mutations of the second kind propagate through the ribosome for considerable distances and are reversed when A-site substrates bind to the ribosome.
Figures
References
-
- Nissen P, Hansen J, Ban N, Moore PB, Steitz TA. The structural basis of ribosome activity in peptide bond synthesis. Science. 2000;289:920–930. - PubMed
-
- Schmeing TM, Seila AC, Hansen JL, Freeborn B, Soukup JK, Scaringe SA, Strobel SA, Moore PB, Steitz TA. A pre-translocational intermediate in protein synthesis observed in crystals of enzymatically active 50S subunits. Nat. Struct. Biol. 2002;9:225–230. - PubMed
-
- Hansen JL, Moore PB, Steitz TA. Structures of five antibiotics bound at the peptidyl transferase center of the large ribosomal subunit. J. Mol. Biol. 2003;330:1061–1075. - PubMed
-
- Schlunzen F, Zarivach R, Harms J, Bashan A, Tocilj A, Albrecht R, Yonath A, Franceschi F. Structural basis for the interaction of antibiotics with the peptidyl transferase centre in eubacteria. Nature. 2001;413:814–821. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
