

Review
doi: 10.1128/JB.00486-08.
Epub 2008 May 2.
Cell-cell communication in bacteria: united we stand
Affiliations
- PMID: 18456806
- PMCID: PMC2446813
- DOI: 10.1128/JB.00486-08
Item in Clipboard
Review
Cell-cell communication in bacteria: united we stand
J Bacteriol.
2008 Jul.
No abstract available
Figures
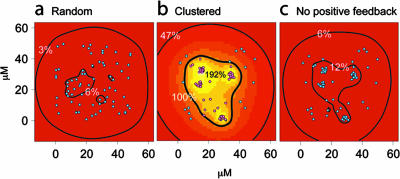
Effect of spatial clustering on QS signaling. A mathematical model for autoinducer systems with or without positive feedback was developed. The model was used to investigate the effect of the spatial arrangement of autoinducer-producing cells on the accumulation of local autoinducer. Comparing a random (a) with a clustered (b) arrangement of the same number of cells, the threshold concentration for induction is reached only within and near the clusters. The figure shows bacterial cells that are not induced in cyan and those that are induced in purple. Comparing the same clustered pattern with (b) and without (c) positive feedback demonstrates that this characteristic of autoinducer production is critical for reaching sufficient autoinducer concentrations for cells to induce autoinducer production and autoinducer-dependent genes. The autoinducer concentration, as a percentage of the threshold concentration, is indicated by contour lines and background color, for which a linear color map from red (<16%) to white (>200%) was used. In panel b, the thick contour line separates the noninduced cells from the induced cells. The three-dimensional domain is viewed from the top, onto an impermeable surface at the bottom. As the domain is otherwise infinite, the autoinducer can diffuse away.
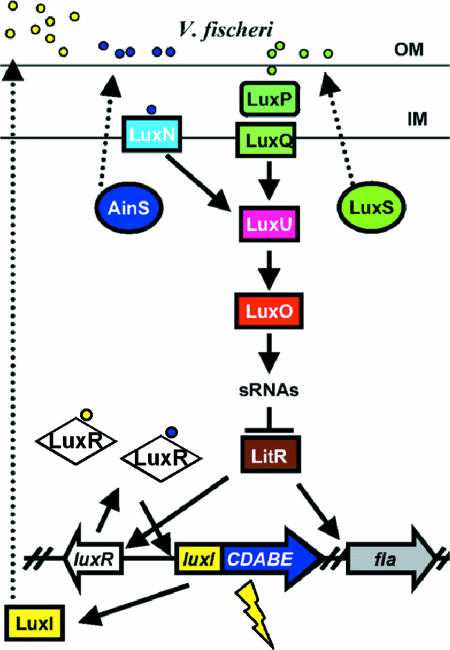
The V. fischeri lux paradigm regulatory cascade. V. fischeri produces three QS signals: the LuxI-produced 3-oxo-hexanoyl-HSL (yellow circles), AinS-produced C8-HSL (blue circles), and a LuxS-produced signal, presumably a furanosyl borate diester (AI-2; green circles). LuxR acts as a receptor for both C8-HSL and 3-O-C6-HSL (61). C8-HSL is also a signal for the membrane-bound LuxN sensor kinase, while AI-2 interacts with the LuxP periplasmic binding protein and the LuxQ sensor kinase. At low cell density (low signal concentrations), LuxN and LuxQ autophosphorylate and transfer the phosphate to LuxU, which in turn phosphorylates the LuxO transcriptional activator. Phosphorylated LuxO is predicted to activate the transcription of sRNAs, which inhibit the production of LitR protein. Increasing population density and signal concentrations switch the kinases to phosphatases, leading to dephosphorylated LuxO and production of LitR. LitR activates luxR and flagellar genes. In turn, LuxR and inducing levels of the 3-O-C6-HSL stimulate light production. (Note that aspects of this model remain to be tested in V. fischeri and are based on the experimentally defined V. harveyi model [102].) OM, outer membrane; IM, inner membrane. (Reprinted from reference with permission.)
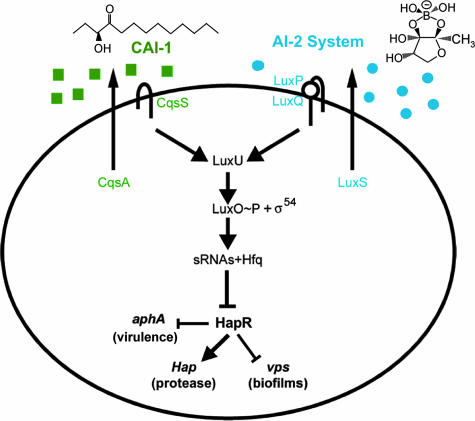
The Vibrio cholerae autoinducer signaling network. The signal synthases CqsA and LuxS produce autoinducers CAI-1 [(S)-3-hydroxytridecan-4-one] and AI-2 [(2S,4S)-2-methyl-2,3,3,4-tetrahydroxytetrahydrofuran borate], respectively. Signal inputs are transduced via LuxO to control levels of HapR. At low cell density, in the absence of autoinducers, hapR expression is repressed, thereby permitting the expression of virulence factors and biofilm formation. At high cell density and in the presence of autoinducers, LuxO is inactive, permitting HapR production. HapR represses virulence and biofilm formation while activating hap protease expression. (Reprinted from reference with permission of the publisher.)
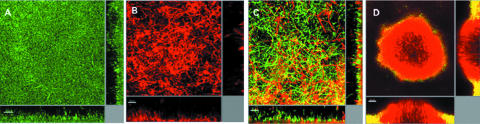
DSF from Stenotrophomonas maltophilia influences biofilm architecture of Pseudomonas aeruginosa PAO1, which does not produce the signal. Images are of 4-day-old biofilms in flow cells. (A) P. aeruginosa PAO1; (B) S. maltophilia K279a; (C) mixed culture of P. aeruginosa PAO1 and S. maltophilia K279a; (D) mixed culture of P. aeruginosa PAO1 and S. maltophilia K279arpfF (DSF negative). Bars, 20 μm. (Courtesy of Max Dow, National University of Ireland, Cork; reproduced with permission).
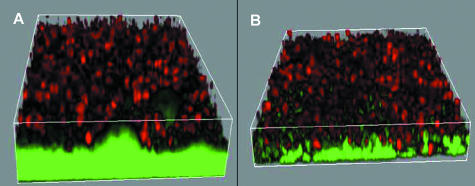
QS-dependent tolerance of P. aeruginosa biofilms toward PMNs. Three-dimensional projection of biofilms (bacteria; green fluorescent) with topical application of PMNs (red fluorescent) on the wild-type strain (A) and the ΔlasR/rhlR mutant (B). (Reprinted from reference with permission of the publisher.)
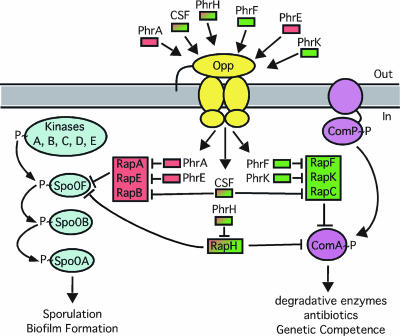
Response to Phr signaling peptides that regulate B. subtilis transcription factors Spo0A and ComA. Phr peptides are secreted pentapeptides that are transported into the cell by an oligopeptide permease (Opp). The peptides interact with cognate Rap proteins to inhibit their phosphatase activity, leading to Spo0F phosphorylation or inhibition of ComA DNA binding activity. Rap/Phr pairs shown in red regulate Spo0A, and Rap/Phr pairs shown in green regulate ComA; red to green shading indicates Phr or Rap interacting with both the Spo0A and ComA pathways. (Reprinted from reference with permission of the publisher.)
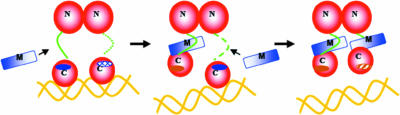
Mechanism of TraMNGR inhibition of TraRNGR, depicting the proposed stepwise dissociation of TraRNGR-DNA by TraMNGR. The first TraMNGR protomer is thought to interact with the exposed linker (green) and TraR C-terminal binding site (dark blue solid oval), leading to the first TraRNGR protomer/DNA dissociation. This interaction repositions the linker and buried TraMNGR binding site (hatched blue oval) of the second TraRNGR protomer so that the next TraMNGR can bind and induce complete TraRNGR/DNA dissociation. (Reprinted from reference with permission of the publisher.)
References
-
- Andrade, M. O., M. C. Alegria, C. R. Guzzo, C. Docena, M. C. Rosa, C. H. Ramos, and C. S. Farah. 2006. The HD-GYP domain of RpfG mediates a direct linkage between the Rpf quorum-sensing pathway and a subset of diguanylate cyclase proteins in the phytopathogen Xanthomonas axonopodis pv citri. Mol. Microbiol. 62537-551. - PubMed
-
- Atkinson, S., T. J. Throup, G. S. Stewart, and P. Williams. 1999. A hierarchical quorum-sensing system in Yersinia pseudotuberculosis is involved in the regulation of motility and clumping. Mol. Microbiol. 331267-1277. - PubMed
-
- Barnard, A. M. L., and G. P. C. Salmond. 2007. Quorum sensing in Erwinia species. Anal. Bioanal. Chem. 387415-424. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
