

Temperature sensitivity and cell division defects in an Escherichia coli strain with mutations in yghB and yqjA, encoding related and conserved inner membrane proteins
- PMID: 18456815
- PMCID: PMC2446817
- DOI: 10.1128/JB.00414-08
Temperature sensitivity and cell division defects in an Escherichia coli strain with mutations in yghB and yqjA, encoding related and conserved inner membrane proteins
Abstract
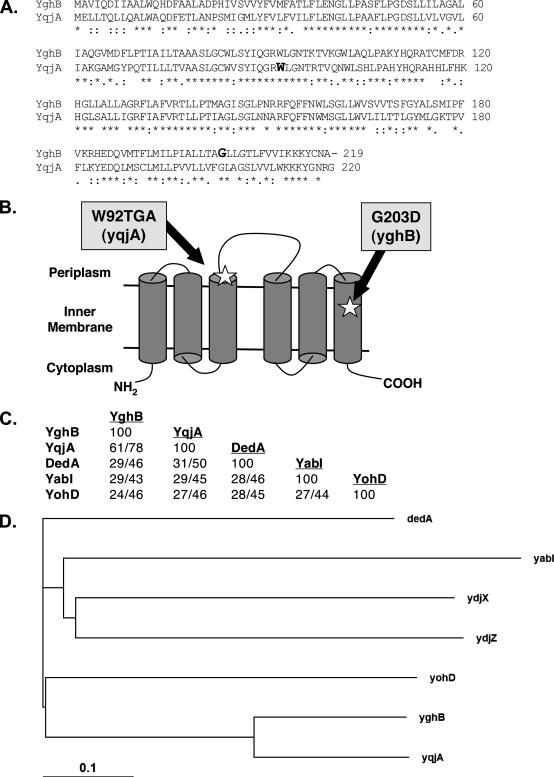
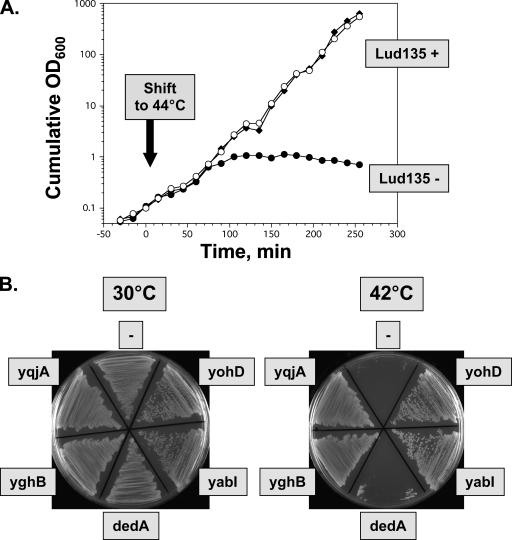
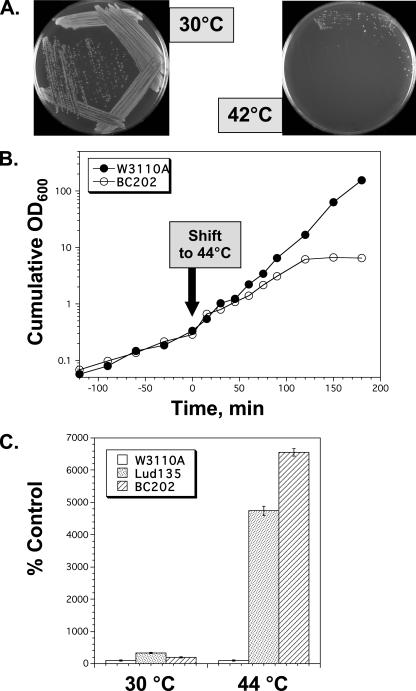
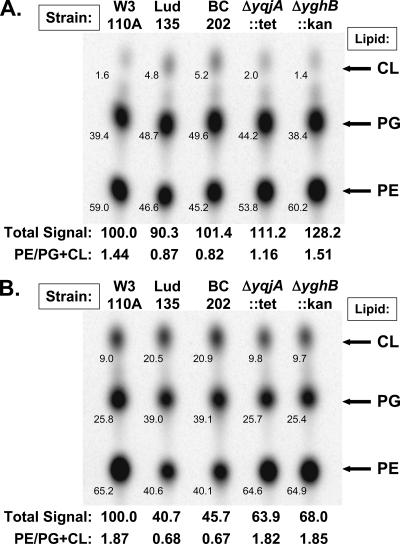
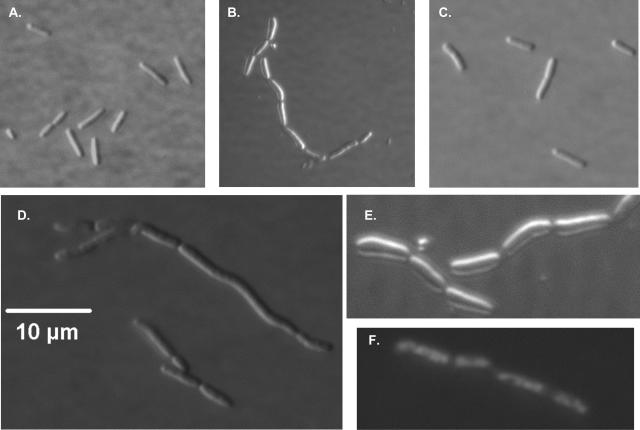
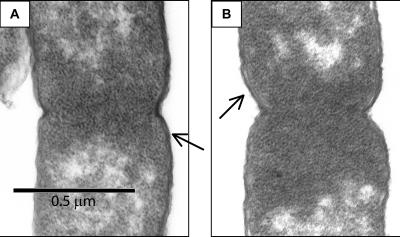
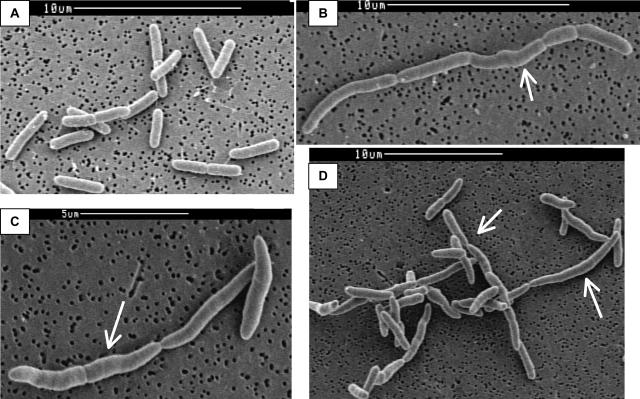
Ludox density gradients were used to enrich for Escherichia coli mutants with conditional growth defects and alterations in membrane composition. A temperature-sensitive mutant named Lud135 was isolated with mutations in two related, nonessential genes: yghB and yqjA. yghB harbors a single missense mutation (G203D) and yqjA contains a nonsense mutation (W92TGA) in Lud135. Both mutations are required for the temperature-sensitive phenotype: targeted deletion of both genes in a wild-type background results in a strain with a similar phenotype and expression of either gene from a plasmid restores growth at elevated temperatures. The mutant has altered membrane phospholipid levels, with elevated levels of acidic phospholipids, when grown under permissive conditions. Growth of Lud135 under nonpermissive conditions is restored by the presence of millimolar concentrations of divalent cations Ca(2+), Ba(2+), Sr(2+), or Mg(2+) or 300 to 500 mM NaCl but not 400 mM sucrose. Microscopic analysis of Lud135 demonstrates a dramatic defect at a late stage of cell division when cells are grown under permissive conditions. yghB and yqjA belong to the conserved and widely distributed dedA gene family, for which no function has been reported. The two open reading frames encode predicted polytopic inner membrane proteins with 61% amino acid identity. It is likely that YghB and YqjA play redundant but critical roles in membrane biology that are essential for completion of cell division in E. coli.
Figures
References
-
- Barr, F. A., and U. Gruneberg. 2007. Cytokinesis: placing and making the final cut. Cell 131847-860. - PubMed
-
- Blattner, F. R., G. Plunkett III, C. A. Bloch, N. T. Perna, V. Burland, M. Riley, J. Collado-Vides, J. D. Glasner, C. K. Rode, G. F. Mayhew, J. Gregor, N. W. Davis, H. A. Kirkpatrick, M. A. Goeden, D. J. Rose, B. Mau, and Y. Shao. 1997. The complete genome sequence of Escherichia coli K-12. Science 2771453-1474. - PubMed
-
- Butland, G., J. M. Peregrin-Alvarez, J. Li, W. Yang, X. Yang, V. Canadien, A. Starostine, D. Richards, B. Beattie, N. Krogan, M. Davey, J. Parkinson, J. Greenblatt, and A. Emili. 2005. Interaction network containing conserved and essential protein complexes in Escherichia coli. Nature 433531-537. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
