

Dynamics of the preprotein translocation channel of the outer membrane of mitochondria
- PMID: 18456827
- PMCID: PMC2479589
- DOI: 10.1529/biophysj.108.131003
Dynamics of the preprotein translocation channel of the outer membrane of mitochondria
Abstract
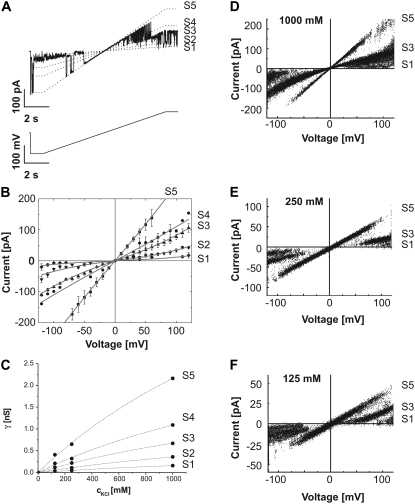
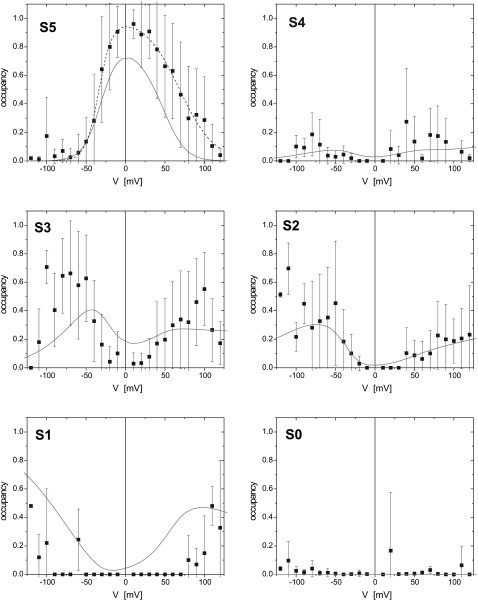
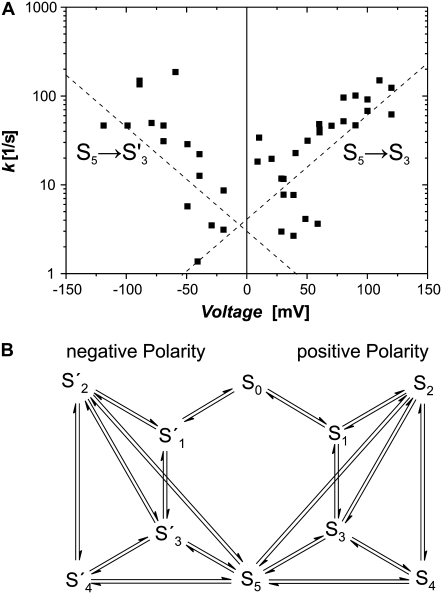
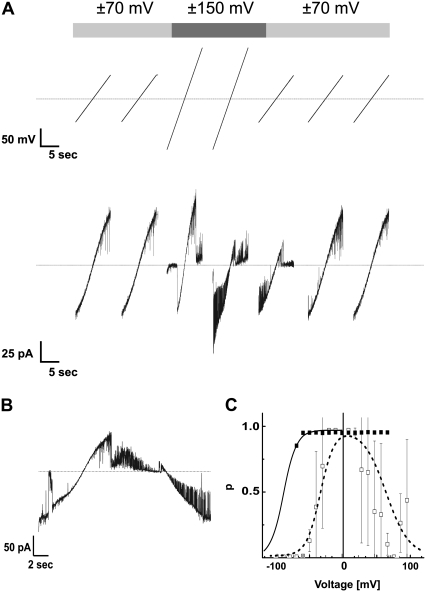
The protein translocase of the outer mitochondrial membrane (TOM) serves as the main entry site for virtually all mitochondrial proteins. Like many other protein translocases it also has an ion channel activity that can be used to study the dynamical properties of this supramolecular complex. We have purified TOM core complex and Tom40, the main pore forming subunit, from mitochondria of the filamentous fungus Neurospora crassa and incorporated them into planar lipid bilayers. We then examined their single channel properties to provide a detailed description of the conformational dynamics of this channel in the absence of its protein substrate. For isolated TOM core complex we have found at least six conductance states. Transitions between these states were voltage-dependent with a bell-shaped open probability distribution and distinct kinetics depending on the polarity of the applied voltage. The states with the largest conductance followed an Ohmic I/V characteristic consistent with a large cylindrical pore with very little interaction with the permeating ions. For the lower conductance states, however, we have observed inverted S-shaped nonlinear current-voltage curves reminiscent to those of much narrower pores where the permeating ions have to surmount an electrostatic energy barrier. At low voltages (<+/-70 mV), purified Tom40 protein did not show any transitions between its conductance states. Prolonged exposure to higher voltages induced similar gating behavior to what we observed for TOM core complex. This effect was time-dependent and reversible, indicating that Tom40 forms not only the pore but also contains the "gating machinery" of the complex. However, for proper functioning, additional proteins (Tom22, Tom7, Tom6, and Tom5) are required that act as a modulator of the pore dynamics by significantly reducing the energy barrier between different conformational states.
Figures



Similar articles
-
Protein import channel of the outer mitochondrial membrane: a highly stable Tom40-Tom22 core structure differentially interacts with preproteins, small tom proteins, and import receptors.Mol Cell Biol. 2001 Apr;21(7):2337-48. doi: 10.1128/MCB.21.7.2337-2348.2001. Mol Cell Biol. 2001. PMID: 11259583 Free PMC article.
-
Tom40, the pore-forming component of the protein-conducting TOM channel in the outer membrane of mitochondria.J Cell Biol. 2001 Jun 11;153(6):1151-60. doi: 10.1083/jcb.153.6.1151. J Cell Biol. 2001. PMID: 11402060 Free PMC article.
-
Identification of Tom5 and Tom6 in the preprotein translocase complex of human mitochondrial outer membrane.Biochem Biophys Res Commun. 2008 May 9;369(3):958-63. doi: 10.1016/j.bbrc.2008.02.150. Epub 2008 Mar 10. Biochem Biophys Res Commun. 2008. PMID: 18331822
-
The transport machinery for the import of preproteins across the outer mitochondrial membrane.Int J Biochem Cell Biol. 2000 Jan;32(1):13-21. doi: 10.1016/s1357-2725(99)00114-4. Int J Biochem Cell Biol. 2000. PMID: 10661891 Review.
-
How does the TOM complex mediate insertion of precursor proteins into the mitochondrial outer membrane?J Cell Biol. 2005 Nov 7;171(3):419-23. doi: 10.1083/jcb.200507147. Epub 2005 Oct 31. J Cell Biol. 2005. PMID: 16260501 Free PMC article. Review.
Cited by
-
Mitochondrial ion channels as therapeutic targets.FEBS Lett. 2010 May 17;584(10):2142-52. doi: 10.1016/j.febslet.2010.02.046. Epub 2010 Feb 20. FEBS Lett. 2010. PMID: 20178788 Free PMC article. Review.
-
Evidence of Distinct Channel Conformations and Substrate Binding Affinities for the Mitochondrial Outer Membrane Protein Translocase Pore Tom40.J Biol Chem. 2015 Oct 23;290(43):26204-17. doi: 10.1074/jbc.M115.642173. Epub 2015 Sep 2. J Biol Chem. 2015. PMID: 26336107 Free PMC article.
-
New insights into the structure and dynamics of the TOM complex in mitochondria.Biochem Soc Trans. 2024 Apr 24;52(2):911-922. doi: 10.1042/BST20231236. Biochem Soc Trans. 2024. PMID: 38629718 Free PMC article. Review.
-
Identification of new channels by systematic analysis of the mitochondrial outer membrane.J Cell Biol. 2017 Nov 6;216(11):3485-3495. doi: 10.1083/jcb.201706043. Epub 2017 Sep 15. J Cell Biol. 2017. PMID: 28916712 Free PMC article.
-
Reconstitution of the membrane protein OmpF into biomimetic block copolymer-phospholipid hybrid membranes.Beilstein J Nanotechnol. 2016 Jun 21;7:881-92. doi: 10.3762/bjnano.7.80. eCollection 2016. Beilstein J Nanotechnol. 2016. PMID: 27547605 Free PMC article.
References
-
- Braun, H. P., and U. K. Schmitz. 1999. The protein-import apparatus of plant mitochondria. Planta. 209:267–274. - PubMed
-
- Lithgow, T. 2000. Targeting of proteins to mitochondria. FEBS Lett. 476:22–26. - PubMed
-
- Pfanner, N., and A. Geissler. 2001. Versatility of the mitochondrial protein import machinery. Nat. Rev. Mol. Cell Biol. 2:339–349. - PubMed
-
- Pfanner, N., N. Wiedemann, C. Meisinger, and T. Lithgow. 2004. Assembling the mitochondrial outer membrane. Nat. Struct. Mol. Biol. 11:1044–1048. - PubMed
-
- Rehling, P., K. Brandner, and N. Pfanner. 2004. Mitochondrial import and the twin-pore translocase. Nat. Rev. Mol. Cell Biol. 5:519–530. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
