

The extracellular metalloprotease of Vibrio tubiashii is a major virulence factor for pacific oyster (Crassostrea gigas) larvae
- PMID: 18456850
- PMCID: PMC2446533
- DOI: 10.1128/AEM.00061-08
The extracellular metalloprotease of Vibrio tubiashii is a major virulence factor for pacific oyster (Crassostrea gigas) larvae
Abstract
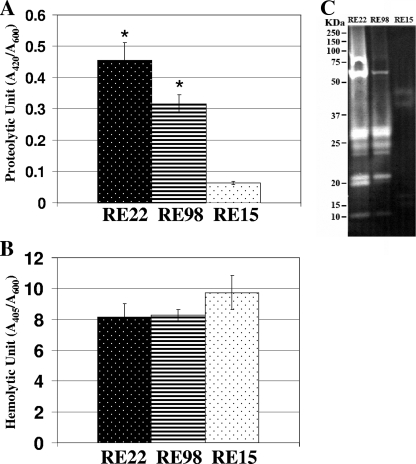
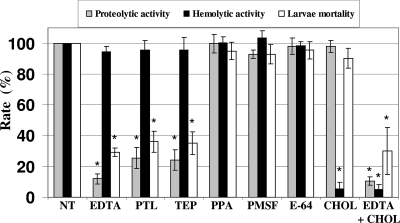
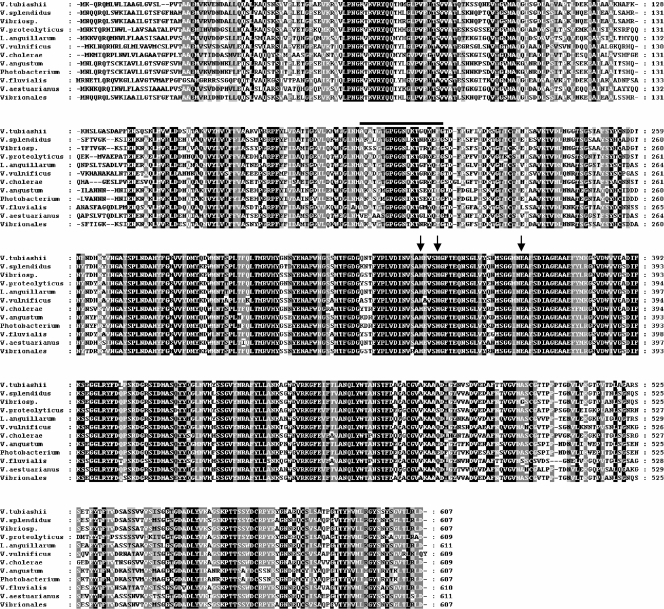
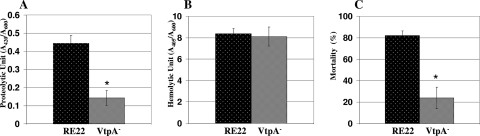
Vibrio tubiashii is a recently reemerging pathogen of larval bivalve mollusks, causing both toxigenic and invasive disease. Marine Vibrio spp. produce an array of extracellular products as potential pathogenicity factors. Culture supernatants of V. tubiashii have been shown to be toxic to oyster larvae and were reported to contain a metalloprotease and a cytolysin/hemolysin. However, the structural genes responsible for these proteins have yet to be identified, and it is uncertain which extracellular products play a role in pathogenicity. We investigated the effects of the metalloprotease and hemolysin secreted by V. tubiashii on its ability to kill Pacific oyster (Crassostrea gigas) larvae. While V. tubiashii supernatants treated with metalloprotease inhibitors severely reduced the toxicity to oyster larvae, inhibition of the hemolytic activity did not affect larval toxicity. We identified structural genes of V. tubiashii encoding a metalloprotease (vtpA) and a hemolysin (vthA). Sequence analyses revealed that VtpA shared high homology with metalloproteases from a variety of Vibrio species, while VthA showed high homology only to the cytolysin/hemolysin of Vibrio vulnificus. Compared to the wild-type strain, a VtpA mutant of V. tubiashii not only produced reduced amounts of protease but also showed decreased toxicity to C. gigas larvae. Vibrio cholerae strains carrying the vtpA or vthA gene successfully secreted the heterologous protein. Culture supernatants of V. cholerae carrying vtpA but not vthA were highly toxic to Pacific oyster larvae. Together, these results suggest that the V. tubiashii extracellular metalloprotease is important in its pathogenicity to C. gigas larvae.
Figures



Similar articles
-
TetR-type transcriptional regulator VtpR functions as a global regulator in Vibrio tubiashii.Appl Environ Microbiol. 2009 Dec;75(24):7602-9. doi: 10.1128/AEM.01016-09. Epub 2009 Oct 16. Appl Environ Microbiol. 2009. PMID: 19837838 Free PMC article.
-
Virulence of metalloproteases produced by Vibrio species on Pacific oyster Crassostrea gigas larvae.Dis Aquat Organ. 2009 Jun 10;85(2):123-31. doi: 10.3354/dao02070. Dis Aquat Organ. 2009. PMID: 19694172
-
The extracellular metalloprotease of Vibrio tubiashii directly inhibits its extracellular haemolysin.Microbiology (Reading). 2009 Jul;155(Pt 7):2296-2305. doi: 10.1099/mic.0.028605-0. Epub 2009 Apr 23. Microbiology (Reading). 2009. PMID: 19389767
-
Regulation systems of protease and hemolysin production in Vibrio vulnificus.Microbiol Immunol. 2017 Jan;61(1):1-11. doi: 10.1111/1348-0421.12465. Microbiol Immunol. 2017. PMID: 28111826 Review.
-
The extracellular proteases produced by Vibrio parahaemolyticus.World J Microbiol Biotechnol. 2018 May 11;34(5):68. doi: 10.1007/s11274-018-2453-4. World J Microbiol Biotechnol. 2018. PMID: 29752585 Review.
Cited by
-
Increased seawater temperature increases the abundance and alters the structure of natural Vibrio populations associated with the coral Pocillopora damicornis.Front Microbiol. 2015 May 18;6:432. doi: 10.3389/fmicb.2015.00432. eCollection 2015. Front Microbiol. 2015. PMID: 26042096 Free PMC article.
-
Insight into the resilience and susceptibility of marine bacteria to T6SS attack by Vibrio cholerae and Vibrio coralliilyticus.PLoS One. 2020 Jan 28;15(1):e0227864. doi: 10.1371/journal.pone.0227864. eCollection 2020. PLoS One. 2020. PMID: 31990915 Free PMC article.
-
Fitness factors in vibrios: a mini-review.Microb Ecol. 2013 May;65(4):826-51. doi: 10.1007/s00248-012-0168-x. Epub 2013 Jan 10. Microb Ecol. 2013. PMID: 23306394 Review.
-
Bacterial community in Sinonovacula constricta intestine and its relationship with culture environment.Appl Microbiol Biotechnol. 2022 Aug;106(13-16):5211-5220. doi: 10.1007/s00253-022-12048-0. Epub 2022 Jul 4. Appl Microbiol Biotechnol. 2022. PMID: 35781839
-
Mechanistic molecular responses of the giant clam Tridacna crocea to Vibrio coralliilyticus challenge.PLoS One. 2020 Apr 10;15(4):e0231399. doi: 10.1371/journal.pone.0231399. eCollection 2020. PLoS One. 2020. PMID: 32276269 Free PMC article.
References
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. S. Truhl. 1991. Current protocols in molecular biology. John Wiley and Sons, New York, NY.
-
- Brown, C. 1981. A study of two shellfish-pathogenic Vibrio strains isolated from a Long Island hatchery during a recent outbreak of disease. J. Shellfish Res. 1:83-87.
-
- Chen, F. R., P. C. Liu, and K. K. Lee. 2000. Lethal attribute of serine protease secreted by Vibrio alginolyticus strains in kuruma prawn Penaeus japonicus. Z. Naturforsch. Sect. C 55:94-99. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
