

A systematic analysis of intronic sequences downstream of 5' splice sites reveals a widespread role for U-rich motifs and TIA1/TIAL1 proteins in alternative splicing regulation
- PMID: 18456862
- PMCID: PMC2493427
- DOI: 10.1101/gr.073155.107
A systematic analysis of intronic sequences downstream of 5' splice sites reveals a widespread role for U-rich motifs and TIA1/TIAL1 proteins in alternative splicing regulation
Abstract
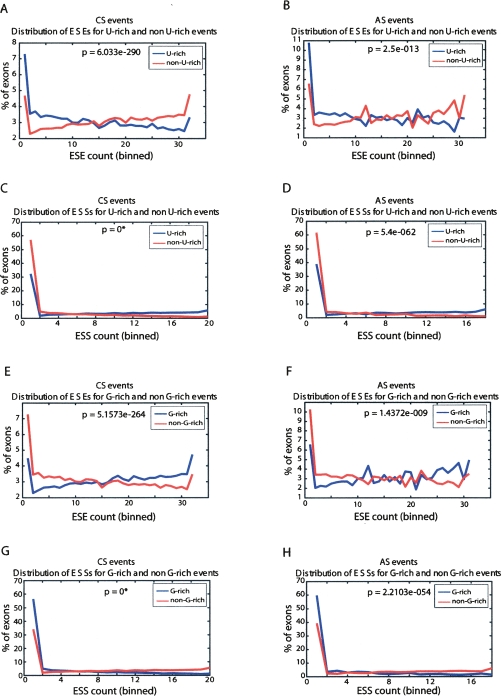
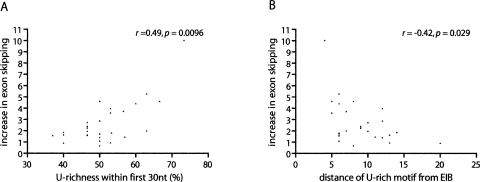
To identify human intronic sequences associated with 5' splice site recognition, we performed a systematic search for motifs enriched in introns downstream of both constitutive and alternative cassette exons. Significant enrichment was observed for U-rich motifs within 100 nucleotides downstream of 5' splice sites of both classes of exons, with the highest enrichment between positions +6 and +30. Exons adjacent to U-rich intronic motifs contain lower frequencies of exonic splicing enhancers and higher frequencies of exonic splicing silencers, compared with exons not followed by U-rich intronic motifs. These findings motivated us to explore the possibility of a widespread role for U-rich motifs in promoting exon inclusion. Since cytotoxic granule-associated RNA binding protein (TIA1) and TIA1-like 1 (TIAL1; also known as TIAR) were previously shown in vitro to bind to U-rich motifs downstream of 5' splice sites, and to facilitate 5' splice site recognition in vitro and in vivo, we investigated whether these factors function more generally in the regulation of splicing of exons followed by U-rich intronic motifs. Simultaneous knockdown of TIA1 and TIAL1 resulted in increased skipping of 36/41 (88%) of alternatively spliced exons associated with U-rich motifs, but did not affect 32/33 (97%) alternatively spliced exons that are not associated with U-rich motifs. The increase in exon skipping correlated with the proximity of the first U-rich motif and the overall "U-richness" of the adjacent intronic region. The majority of the alternative splicing events regulated by TIA1/TIAL1 are conserved in mouse, and the corresponding genes are associated with diverse cellular functions. Based on our results, we estimate that approximately 15% of alternative cassette exons are regulated by TIA1/TIAL1 via U-rich intronic elements.
Figures


References
-
- Ansley S.J., Badano J.L., Blacque O.E., Hill J., Hoskins B.E., Leitch C.C., Kim J.C., Ross A.J., Eichers E.R., Teslovich T.M., Badano J.L., Blacque O.E., Hill J., Hoskins B.E., Leitch C.C., Kim J.C., Ross A.J., Eichers E.R., Teslovich T.M., Blacque O.E., Hill J., Hoskins B.E., Leitch C.C., Kim J.C., Ross A.J., Eichers E.R., Teslovich T.M., Hill J., Hoskins B.E., Leitch C.C., Kim J.C., Ross A.J., Eichers E.R., Teslovich T.M., Hoskins B.E., Leitch C.C., Kim J.C., Ross A.J., Eichers E.R., Teslovich T.M., Leitch C.C., Kim J.C., Ross A.J., Eichers E.R., Teslovich T.M., Kim J.C., Ross A.J., Eichers E.R., Teslovich T.M., Ross A.J., Eichers E.R., Teslovich T.M., Eichers E.R., Teslovich T.M., Teslovich T.M., et al. Basal body dysfunction is a likely cause of pleiotropic Bardet-Biedl syndrome. Nature. 2003;425:628–633. - PubMed
-
- Barash Y., Bejerano G., Friedman N., Bejerano G., Friedman N., Friedman N. A simple hyper-geometric approach for discovering putative transcription factor binding sites. Lect. Notes Comp. Sci. 2001;2149:278–293.
-
- Beck A.R., Medley Q.G., O’Brien S., Anderson P., Streuli M., Medley Q.G., O’Brien S., Anderson P., Streuli M., O’Brien S., Anderson P., Streuli M., Anderson P., Streuli M., Streuli M. Structure, tissue distribution and genomic organization of the murine RRM-type RNA binding proteins TIA1 and TIAL1. Nucleic Acids Res. 1996;24:3829–3835. - PMC - PubMed
-
- Beissbarth T., Speed T.P., Speed T.P. GOstat: Find statistically overrepresented Gene Ontologies within a group of genes. Bioinformatics. 2004;20:1464–1465. - PubMed
-
- Blencowe B.J. Alternative splicing: New insights from global analyses. Cell. 2006;126:37–47. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous