

Human betaine-homocysteine methyltransferase (BHMT) and BHMT2: common gene sequence variation and functional characterization
- PMID: 18457970
- PMCID: PMC2515933
- DOI: 10.1016/j.ymgme.2008.03.013
Human betaine-homocysteine methyltransferase (BHMT) and BHMT2: common gene sequence variation and functional characterization
Abstract
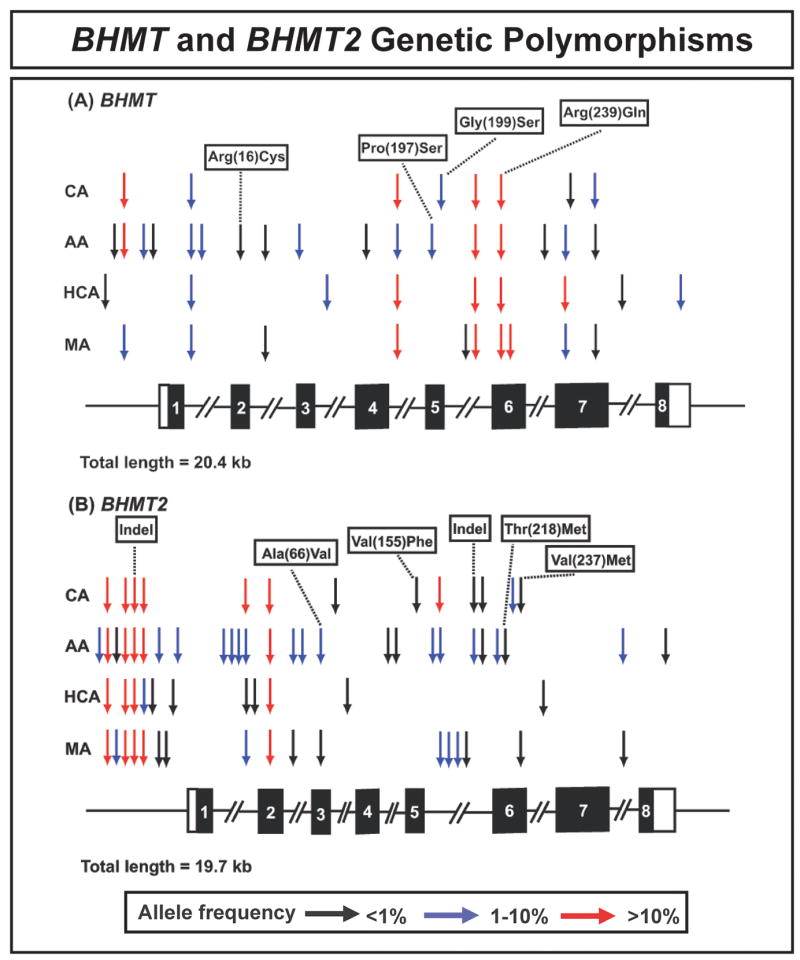
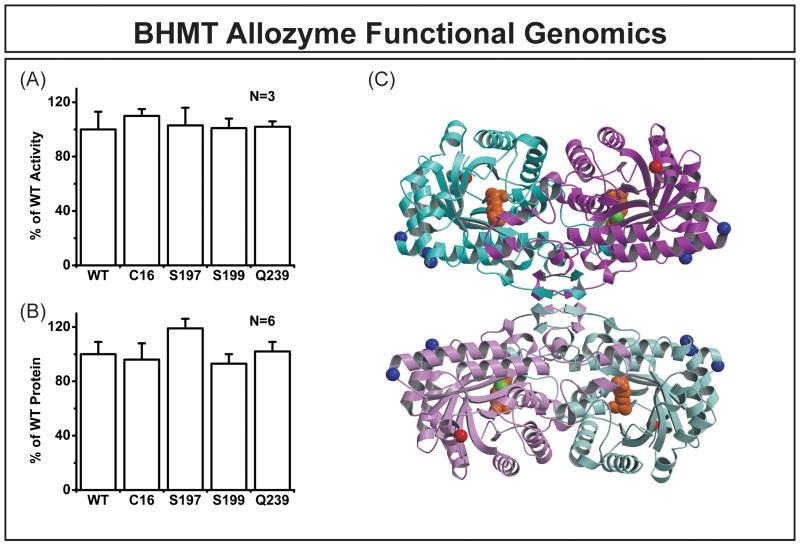
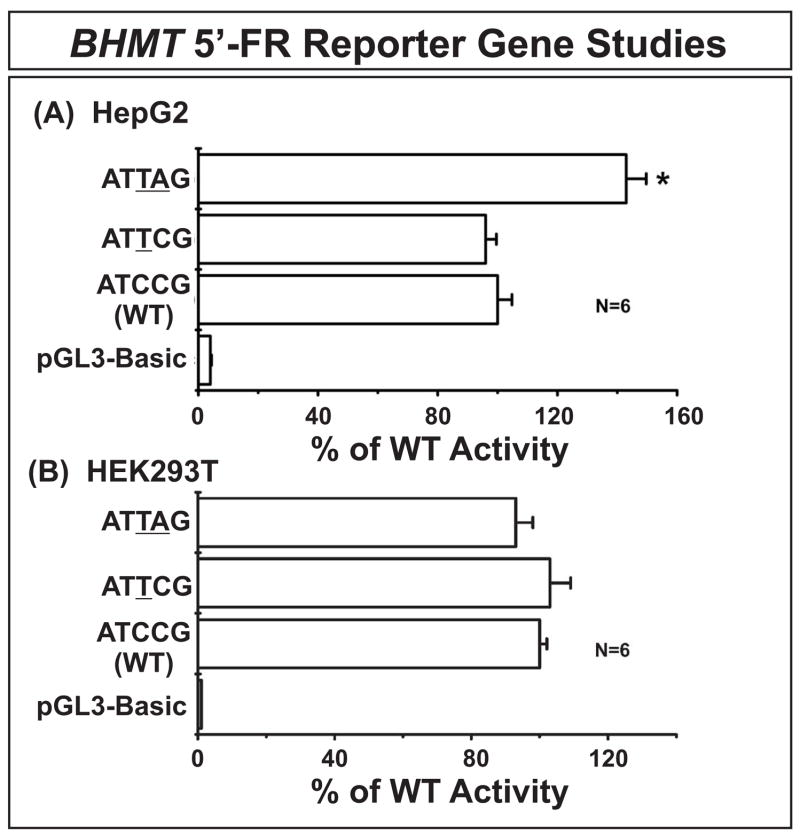
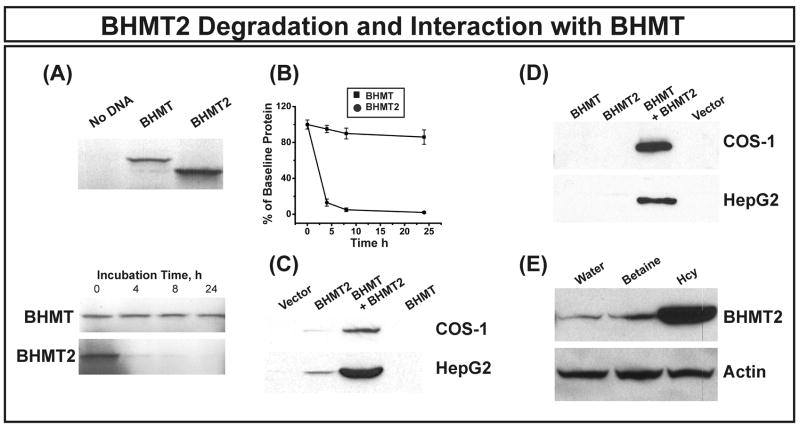
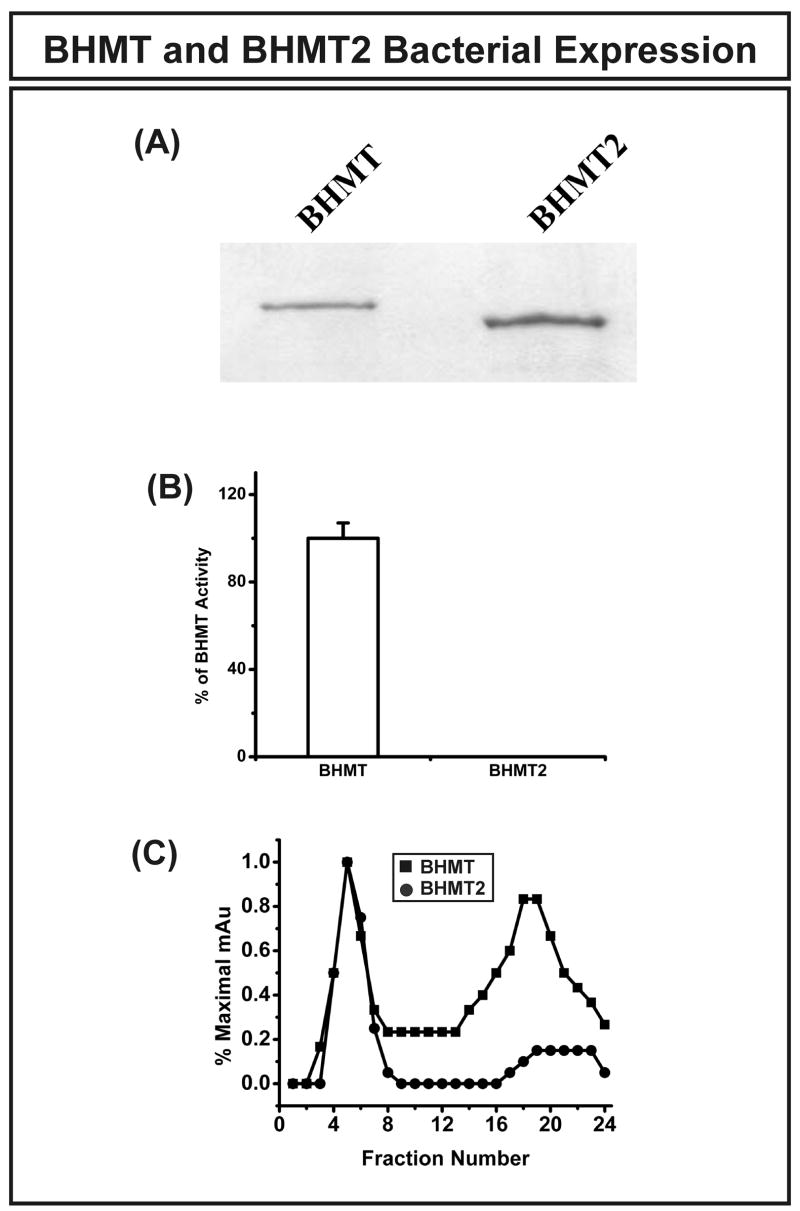
Betaine-homocysteine methyltransferase (BHMT) catalyzes the remethylation of homocysteine. BHMT2 encodes a protein 73% identical in amino acid sequence to BHMT, but the function of BHMT2 remains unclear. We set out to identify and functionally characterize common genetic variation in BHMT and BHMT2. Specifically, we sequenced exons, exon-intron splice junctions and the 5'-flanking regions (5'-FRs) of BHMT and BHMT2 using 240 DNA samples from four ethnic groups. Twenty-five single nucleotide polymorphisms (SNPs), including 4 nonsynonymous SNPs, and 39 SNPs, including 4 nonsynonymous, were observed in BHMT and BHMT2, respectively. BHMT wild type (WT) and variant allozymes were expressed in COS-1 cells. Variant allozymes showed no significant differences from WT in levels of enzyme activity or immunoreactive protein, but there were statistically significant differences in apparent K(m) values. Luciferase reporter gene constructs were created for the three most common BHMT 5'-FR haplotypes, and significant variation was observed in the ability of these constructs to drive transcription. Although BHMT2 mRNA has been observed in human liver and kidney, expression of the protein has not been reported. We were unable to express BHMT2 in mammalian cells, and the protein aggregated after bacterial expression. Furthermore, BHMT2 was rapidly degraded in a rabbit reticulocyte lysate, but it could be stabilized by cotransfection of COS-1 cells with BHMT and, after cotransfection, it coprecipitated with BHMT. These studies have defined common genetic variation in BHMT and BHMT2 and functionally characterized BHMT SNPs. They may also help to explain why BHMT2 has not previously been defined functionally.
Figures

References
-
- Carmel R, Jacobsen DW. Homocysteine in Health and Disease. Cambridge University Press; New York: 2001.
-
- Millian NS, Garrow TA. Human betaine-homocysteine methyltransferase is a zinc metalloenzyme. Arch Biochem Biophys. 1998;356:93–8. - PubMed
-
- Finkelstein JD, Martin JJ. Methionine metabolism in mammals. Distribution of homocysteine between competing pathways. J Biol Chem. 1984;259:9508–13. - PubMed
-
- Sunden SL, Renduchintala MS, Park EI, Miklasz SD, Garrow TA. Betaine-homocysteine methyltransferase expression in porcine and human tissues and chromosomal localization of the human gene. Arch Biochem Biophys. 1997;345:171–4. - PubMed
-
- Park EI, Garrow TA. Interaction between dietary methionine and methyl donor intake on rat liver betaine-homocysteine methyltransferase gene expression and organization of the human gene. J Biol Chem. 1999;274:7816–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
