

Caenorhabditis elegans genes required for the engulfment of apoptotic corpses function in the cytotoxic cell deaths induced by mutations in lin-24 and lin-33
- PMID: 18458102
- PMCID: PMC2390618
- DOI: 10.1534/genetics.108.087221
Caenorhabditis elegans genes required for the engulfment of apoptotic corpses function in the cytotoxic cell deaths induced by mutations in lin-24 and lin-33
Abstract
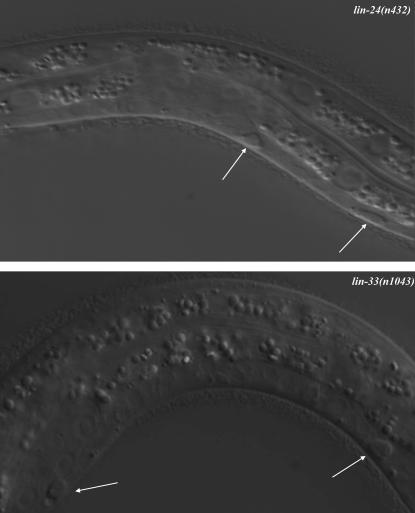
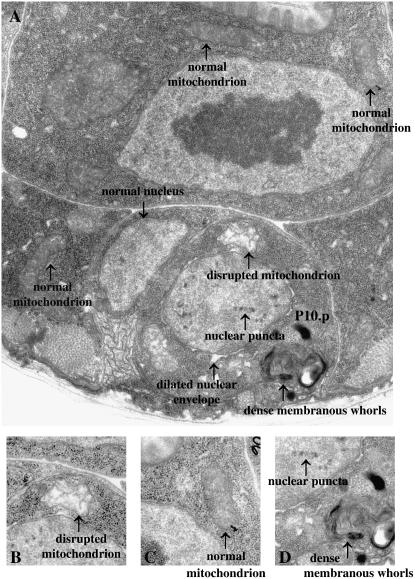
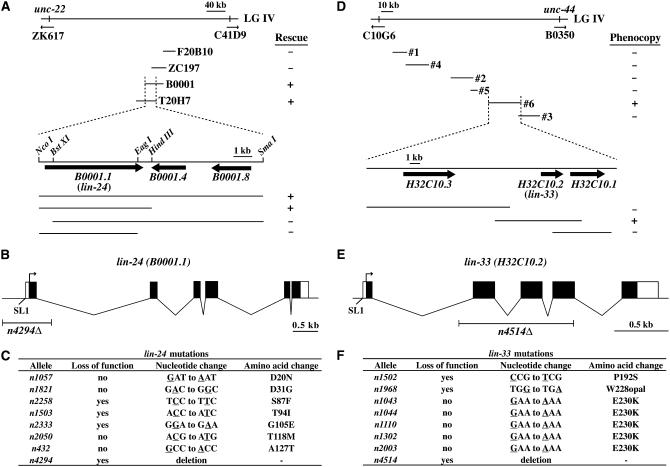
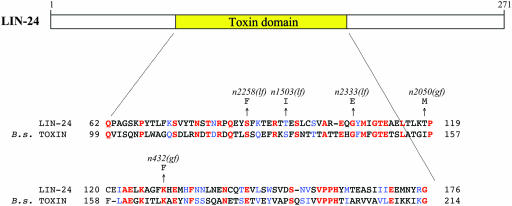
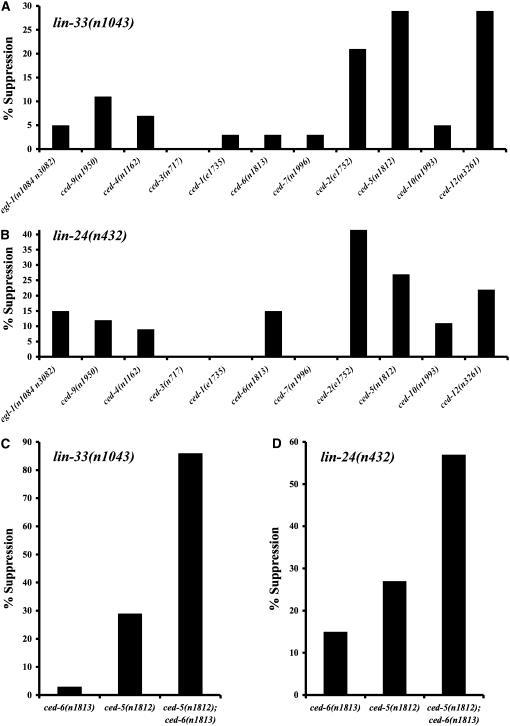
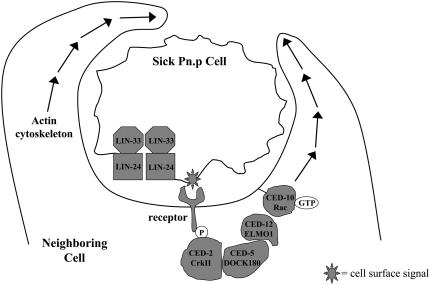
Two types of cell death have been studied extensively in Caenorhabditis elegans, programmed cell death and necrosis. We describe a novel type of cell death that occurs in animals containing mutations in either of two genes, lin-24 and lin-33. Gain-of-function mutations in lin-24 and lin-33 cause the inappropriate deaths of many of the Pn.p hypodermal blast cells and prevent the surviving Pn.p cells from expressing their normal developmental fates. The abnormal Pn.p cells in lin-24 and lin-33 mutant animals are morphologically distinct from the dying cells characteristic of C. elegans programmed cell deaths and necrotic cell deaths. lin-24 encodes a protein with homology to bacterial toxins. lin-33 encodes a novel protein. The cytotoxicity caused by mutation of either gene requires the function of the other. An evolutionarily conserved set of genes required for the efficient engulfment and removal of both apoptotic and necrotic cell corpses is required for the full cell-killing effect of mutant lin-24 and lin-33 genes, suggesting that engulfment promotes these cytotoxic cell deaths.
Figures
Similar articles
-
DPL-1 DP, LIN-35 Rb and EFL-1 E2F act with the MCD-1 zinc-finger protein to promote programmed cell death in Caenorhabditis elegans.Genetics. 2007 Apr;175(4):1719-33. doi: 10.1534/genetics.106.068148. Epub 2007 Jan 21. Genetics. 2007. PMID: 17237514 Free PMC article.
-
The Caenorhabditis elegans gene lin-26 is required to specify the fates of hypodermal cells and encodes a presumptive zinc-finger transcription factor.Development. 1994 Sep;120(9):2359-68. doi: 10.1242/dev.120.9.2359. Development. 1994. PMID: 7956818
-
The C. elegans cell corpse engulfment gene ced-7 encodes a protein similar to ABC transporters.Cell. 1998 Jun 12;93(6):951-60. doi: 10.1016/s0092-8674(00)81201-5. Cell. 1998. PMID: 9635425
-
LIN-35 beyond its classical roles: its function in the stress response.Int J Dev Biol. 2021;65(4-5-6):377-382. doi: 10.1387/ijdb.200194rn. Int J Dev Biol. 2021. PMID: 32930365 Review.
-
Repurposing the Killing Machine: Non-canonical Roles of the Cell Death Apparatus in Caenorhabditis elegans Neurons.Front Cell Dev Biol. 2022 Feb 14;10:825124. doi: 10.3389/fcell.2022.825124. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35237604 Free PMC article. Review.
Cited by
-
Endogenous pore-forming protein complex targets acidic glycosphingolipids in lipid rafts to initiate endolysosome regulation.Commun Biol. 2019 Feb 11;2:59. doi: 10.1038/s42003-019-0304-y. eCollection 2019. Commun Biol. 2019. PMID: 30775460 Free PMC article.
-
Loss of the RhoGAP SRGP-1 promotes the clearance of dead and injured cells in Caenorhabditis elegans.Nat Cell Biol. 2011 Jan;13(1):79-86. doi: 10.1038/ncb2138. Epub 2010 Dec 19. Nat Cell Biol. 2011. PMID: 21170032 Free PMC article.
-
SRGP-1 regulation, targets, and contribution to cell killing in C. elegans.Small GTPases. 2011 May;2(3):177-181. doi: 10.4161/sgtp.2.3.16700. Small GTPases. 2011. PMID: 21776421 Free PMC article.
-
Noncanonical cell death in the nematode Caenorhabditis elegans.Methods Enzymol. 2014;545:157-80. doi: 10.1016/B978-0-12-801430-1.00007-X. Methods Enzymol. 2014. PMID: 25065890 Free PMC article. Review.
-
Integrin α PAT-2/CDC-42 signaling is required for muscle-mediated clearance of apoptotic cells in Caenorhabditis elegans.PLoS Genet. 2012;8(5):e1002663. doi: 10.1371/journal.pgen.1002663. Epub 2012 May 17. PLoS Genet. 2012. PMID: 22615577 Free PMC article.
References
-
- Abraham, M. C., Y. Lu and S. Shaham, 2007. A morphologically conserved nonapoptotic program promotes linker cell death in Caenorhabditis elegans. Dev. Cell. 12 73–86. - PubMed
-
- Akakura, S., B. Kar, S. Singh, L. Cho, N. Tibrewal et al., 2005. C-terminal SH3 domain of CrkII regulates the assembly and function of the DOCK180/ELMO Rac-GEF. J. Cell. Physiol. 204 344–351. - PubMed
-
- Bargmann, C. I., E. Hartwieg and H. R. Horvitz, 1993. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell 74 515–527. - PubMed
-
- Barstead, R. J., and R. H. Waterston, 1989. The basal component of the nematode dense-body is vinculin. J. Biol. Chem. 264 10177–10185. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
