

Pitx2 is functionally important in the early stages of vascular smooth muscle cell differentiation
- PMID: 18458156
- PMCID: PMC2364692
- DOI: 10.1083/jcb.200711145
Pitx2 is functionally important in the early stages of vascular smooth muscle cell differentiation
Abstract
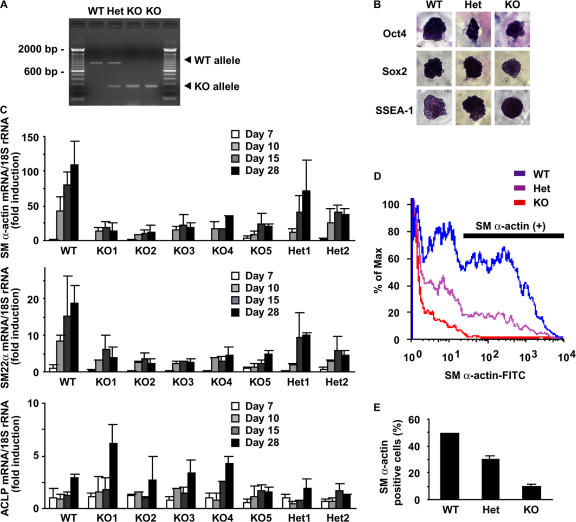
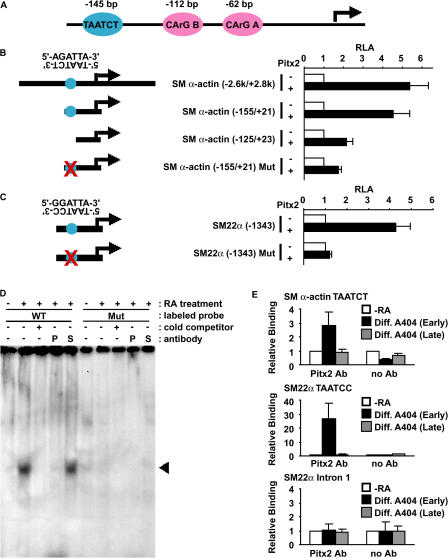
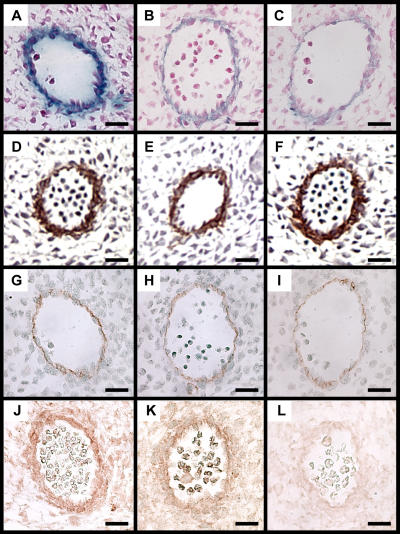
Mechanisms that control vascular smooth muscle cell (SMC) differentiation are poorly understood. We identify Pitx2 as a previously unknown homeodomain transcription factor that is rapidly induced in an in vitro model of SMC differentiation from multipotent stem cells. Pitx2 induces expression of multiple SMC differentiation marker genes by binding to a TAATC(C/T) cis-element, by interacting with serum response factor, and by increasing histone acetylation levels within the promoters of SMC differentiation marker genes. Suppression of Pitx2 reduces expression of SMC differentiation marker genes in the early stages of SMC differentiation in vitro, whereas Prx1, another homeodomain protein, regulates SMC differentiation marker genes in fully differentiated SMCs. Pitx2, but not Prx1, knockout mouse embryos exhibit impaired induction of SMC differentiation markers in the dorsal aorta and branchial arch arteries. Our results demonstrate that Pitx2 functions to regulate the early stages of SMC differentiation.
Figures





References
-
- Bergwerff, M., A.C. Gittenberger-de Groot, L.J. Wisse, M.C. DeRuiter, A. Wessels, J.F. Martin, E.N. Olson, and M.J. Kern. 2000. Loss of function of the Prx1 and Prx2 homeobox genes alters architecture of the great elastic arteries and ductus arteriosus. Virchows Arch. 436:12–19. - PubMed
-
- Carmeliet, P. 2000. Mechanisms of angiogenesis and arteriogenesis. Nat. Med. 6:389–395. - PubMed
-
- Chen, J., C.M. Kitchen, J.W. Streb, and J.M. Miano. 2002. Myocardin: a component of a molecular switch for smooth muscle differentiation. J. Mol. Cell. Cardiol. 34:1345–1356. - PubMed
-
- Drab, M., H. Haller, R. Bychkov, B. Erdmann, C. Lindschau, H. Haase, I. Morano, F.C. Luft, and A.M. Wobus. 1997. From totipotent embryonic stem cells to spontaneously contracting smooth muscle cells: a retinoic acid and db-cAMP in vitro differentiation model. FASEB J. 11:905–915. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
