

Centrosomal RNA correlates with intron-poor nuclear genes in Spisula oocytes
- PMID: 18458332
- PMCID: PMC2383940
- DOI: 10.1073/pnas.0802293105
Centrosomal RNA correlates with intron-poor nuclear genes in Spisula oocytes
Abstract
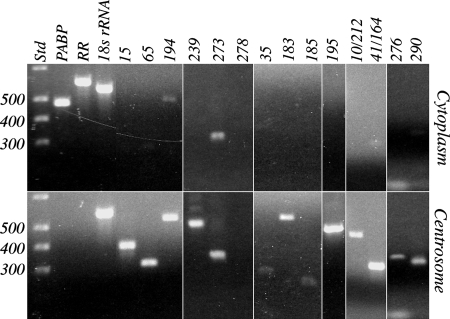
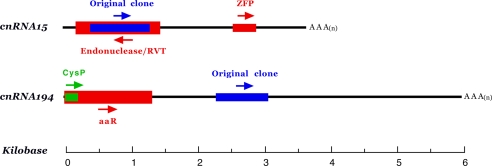
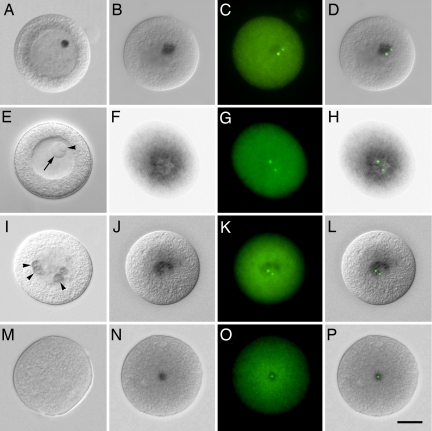
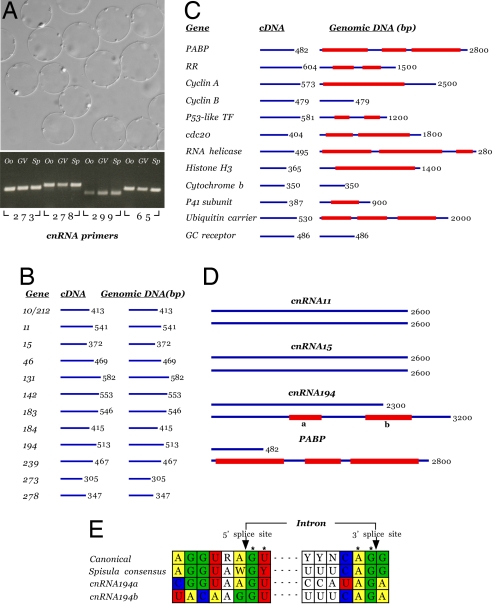
The evolutionary origin of centriole/kinetosomes, centrosomes, and other microtubule organizing centers (MTOCs), whether by direct filiation or symbiogenesis, has been controversial for >50 years. Centrioles, like mitochondria and chloroplasts, duplicate independently of the nucleus and constitute a heritable system independent of chromosomal DNA. Nucleic acids endogenous to the MTOC would support evolutionary origin by symbiogenesis. To date, most reports of centrosome-associated nucleic acids have used generalized reagents such as RNases and nucleic acid dyes. Here, from a library of RNAs extracted from isolated surf clam (Spisula solidissima) centrosomes, we describe a group of centrosome-associated transcripts representing a structurally unique intron-poor collection of nuclear genes skewed toward nucleic acid metabolism. Thus, we resolve the debate over the existence of centrosome-associated RNA (cnRNA). A subset of cnRNAs contain functional domains that are highly conserved across distant taxa, such as nucleotide polymerase motifs. In situ localization of cnRNA65, a molecule with an RNA polymerase domain, showed it is present in the intact oocyte nucleus (germinal vesicle). Its expression, therefore, precedes the appearance of gamma-tubulin-containing centrosomes. At this stage, the in situ signal resembles the nucleolinus, a poorly understood organelle proposed to play a role in spindle formation. After oocyte activation and germinal vesicle breakdown, cnRNA65 persists as a cytoplasmic patch within which gamma-tubulin-stained centrosomes can be seen. These observations provoke the question of whether cnRNAs and the nucleolinus serve as cytological progenitors of the centrosome and may support a symbiogenetic model for its evolution.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Centrosome-associated RNA in surf clam oocytes.Proc Natl Acad Sci U S A. 2006 Jun 13;103(24):9034-8. doi: 10.1073/pnas.0602859103. Epub 2006 Jun 5. Proc Natl Acad Sci U S A. 2006. PMID: 16754862 Free PMC article.
-
Rediscovery of the nucleolinus, a dynamic RNA-rich organelle associated with the nucleolus, spindle, and centrosomes.Proc Natl Acad Sci U S A. 2010 Aug 3;107(31):13718-23. doi: 10.1073/pnas.1008469107. Epub 2010 Jul 19. Proc Natl Acad Sci U S A. 2010. PMID: 20643950 Free PMC article.
-
Composition and dynamics of the nucleolinus, a link between the nucleolus and cell division apparatus in surf clam (Spisula) oocytes.J Biol Chem. 2012 Feb 24;287(9):6702-13. doi: 10.1074/jbc.M111.288506. Epub 2012 Jan 4. J Biol Chem. 2012. PMID: 22219192 Free PMC article.
-
Discovery of centrosomal RNA and centrosomal hypothesis of cellular ageing and differentiation.Nucleosides Nucleotides Nucleic Acids. 2012;31(3):172-83. doi: 10.1080/15257770.2011.648362. Nucleosides Nucleotides Nucleic Acids. 2012. PMID: 22356233 Review.
-
The centrosome in Drosophila oocyte development.Curr Top Dev Biol. 2000;49:385-407. doi: 10.1016/s0070-2153(99)49019-2. Curr Top Dev Biol. 2000. PMID: 11005029 Review.
Cited by
-
RNA in centrosomes: structure and possible functions.Protoplasma. 2013 Feb;250(1):397-405. doi: 10.1007/s00709-012-0422-6. Epub 2012 Jun 10. Protoplasma. 2013. PMID: 22684578
-
The Emerging Role of ncRNAs and RNA-Binding Proteins in Mitotic Apparatus Formation.Noncoding RNA. 2020 Mar 20;6(1):13. doi: 10.3390/ncrna6010013. Noncoding RNA. 2020. PMID: 32245090 Free PMC article. Review.
-
The Cilioprotist Cytoskeleton , a Model for Understanding How Cell Architecture and Pattern Are Specified: Recent Discoveries from Ciliates and Comparable Model Systems.Methods Mol Biol. 2022;2364:251-295. doi: 10.1007/978-1-0716-1661-1_13. Methods Mol Biol. 2022. PMID: 34542858 Review.
-
centrocortin RNA localization to centrosomes is regulated by FMRP and facilitates error-free mitosis.J Cell Biol. 2020 Dec 7;219(12):e202004101. doi: 10.1083/jcb.202004101. J Cell Biol. 2020. PMID: 33196763 Free PMC article.
-
Centriole evolution.Curr Opin Cell Biol. 2009 Feb;21(1):14-9. doi: 10.1016/j.ceb.2009.01.008. Epub 2009 Feb 3. Curr Opin Cell Biol. 2009. PMID: 19196504 Free PMC article. Review.
References
-
- Chapman MJ, Dolan MF, Margulis L. Centrioles and kinetosomes: Form, function, and evolution. Q Rev Biol. 2000;75:409–429. - PubMed
-
- Chapman MJ, Alliegro MC. A symbiogenetic basis for the centrosome? Symbiosis. 2007;44:23–32.
-
- Bornens M, Azimzadeh J. Origins and Evolution of Eukaryotic Endomembranes and Cytoskeleton. In: Jekely G, editor. Austin, TX: Landes Bioscience; 2007.
-
- Palazzo RE, Vogel JM. Isolation of centrosomes from Spisula solidissima oocytes. Methods Cell Biol. 1999;61:35–56. - PubMed
-
- Montgomery TH. Comparative cytological studies, with especial regard to the morphology of the nucleolus. J Morph. 1899;15:265.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
