

Mitochondrial copper(I) transfer from Cox17 to Sco1 is coupled to electron transfer
- PMID: 18458339
- PMCID: PMC2383975
- DOI: 10.1073/pnas.0800019105
Mitochondrial copper(I) transfer from Cox17 to Sco1 is coupled to electron transfer
Abstract
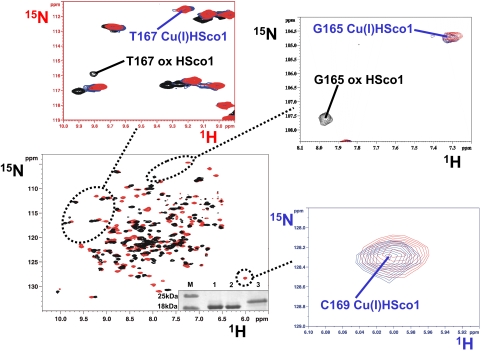
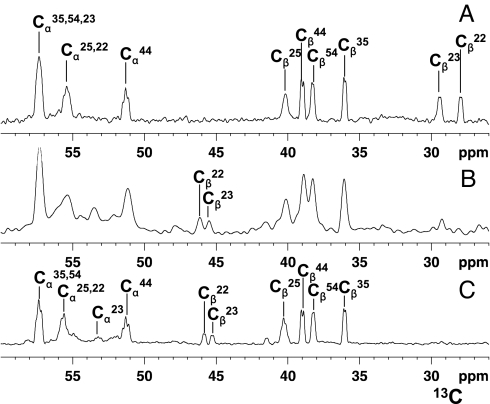
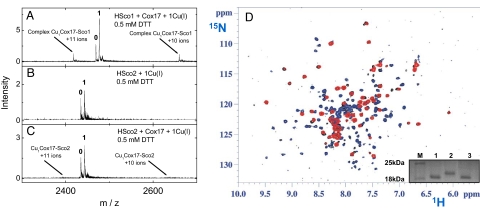
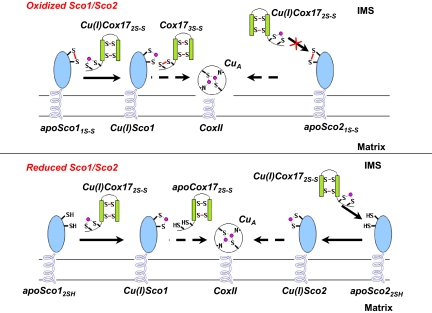
The human protein Cox17 contains three pairs of cysteines. In the mitochondrial intermembrane space (IMS) it exists in a partially oxidized form with two S-S bonds and two reduced cysteines (HCox17(2S-S)). HCox17(2S-S) is involved in copper transfer to the human cochaperones Sco1 and Cox11, which are implicated in the assembly of cytochrome c oxidase. We show here that Cu(I)HCox17(2S-S), i.e., the copper-loaded form of the protein, can transfer simultaneously copper(I) and two electrons to the human cochaperone Sco1 (HSco1) in the oxidized state, i.e., with its metal-binding cysteines forming a disulfide bond. The result is Cu(I)HSco1 and the fully oxidized apoHCox17(3S-S), which can be then reduced by glutathione to apoHCox17(2S-S). The HSco1/HCox17(2S-S) redox reaction is thermodynamically driven by copper transfer. These reactions may occur in vivo because HSco1 can be found in the partially oxidized state within the IMS, consistent with the variable redox properties of the latter compartment. The electron transfer-coupled metallation of HSco1 can be a mechanism within the IMS for an efficient specific transfer of the metal to proteins, where metal-binding thiols are oxidized. The same reaction of copper-electron-coupled transfer does not occur with the human homolog of Sco1, HSco2, for kinetic reasons that may be ascribed to the lack of a specific metal-bridged protein-protein complex, which is instead observed in the Cu(I)HCox17(2S-S)/HSco1 interaction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Finney LA, O'Halloran TV. Transition metal speciation in the cell: Insights from the chemistry of metal ion receptors. Science. 2003;300:931–936. - PubMed
-
- Tsukihara T, et al. Structures of metal sites of oxidized bovine heart cytochrome c oxidase at 2.8 Å. Science. 1995;269:1069–1074. - PubMed
-
- Tsukihara T, et al. The whole structure of the 13-subunit oxidized cytochrome c oxidase at 2.8 Å. Science. 1996;272:1136–1144. - PubMed
-
- Carr HS, Winge DR. Assembly of cytochrome c oxidase within the mitochondrion. Acc Chem Res. 2003;36:309–316. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
