

Inhibition of Wnt signaling by the osteoblast-specific transcription factor Osterix
- PMID: 18458345
- PMCID: PMC2383965
- DOI: 10.1073/pnas.0710831105
Inhibition of Wnt signaling by the osteoblast-specific transcription factor Osterix
Abstract
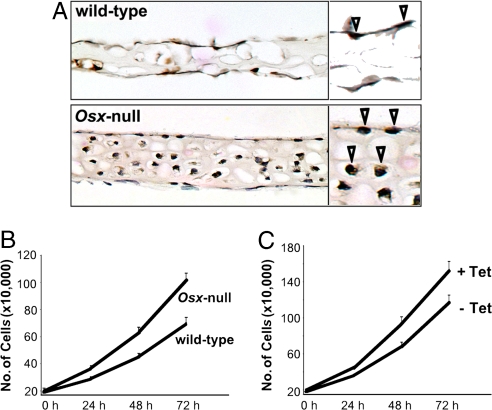
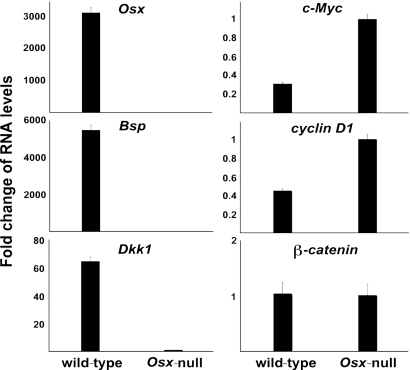
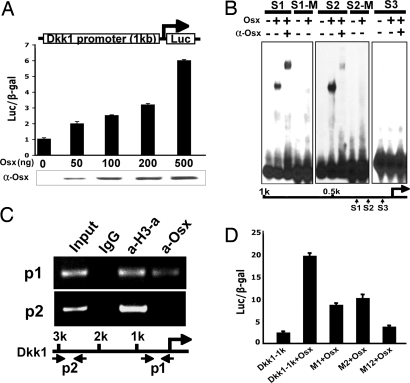
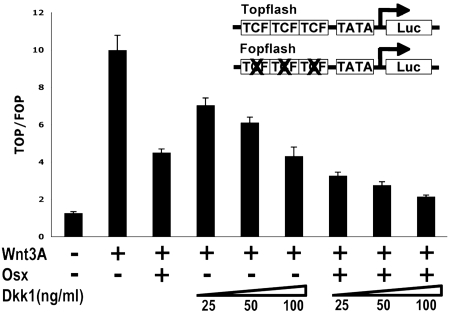
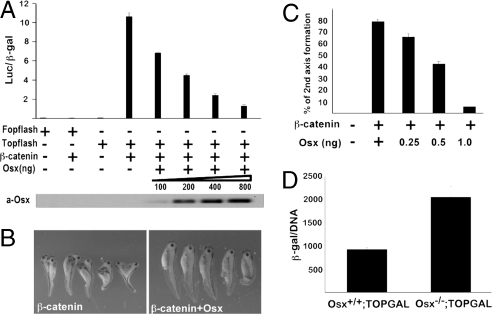
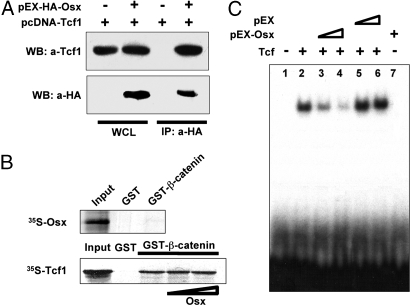
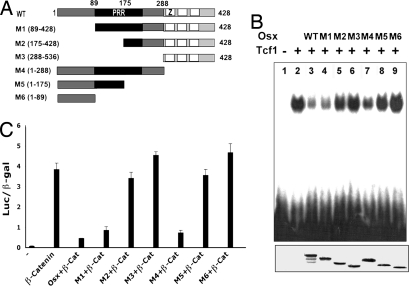
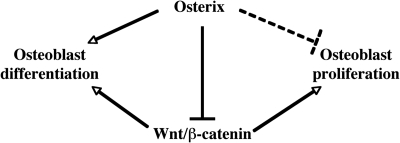
The recent identification of the genes responsible for several human genetic diseases affecting bone homeostasis and the characterization of mouse models for these diseases indicated that canonical Wnt signaling plays a critical role in the control of bone mass. Here, we report that the osteoblast-specific transcription factor Osterix (Osx), which is required for osteoblast differentiation, inhibits Wnt pathway activity. First, in calvarial cells of embryonic day (E)18.5 Osx-null embryos, expression of the Wnt antagonist Dkk1 was abolished, and that of Wnt target genes c-Myc and cyclin D1 was increased. Moreover, our studies demonstrated that Osx bound to and activated the Dkk1 promoter. In addition, Osx inhibited beta-catenin-induced Topflash reporter activity and beta-catenin-induced secondary axis formation in Xenopus embryos. Importantly, in calvaria of E18.5 Osx-null embryos harboring the TOPGAL reporter transgene, beta-galactosidase activity was increased, suggesting that Osx inhibited the Wnt pathway in osteoblasts in vivo. Our data further showed that Osx disrupted binding of Tcf to DNA, providing a likely mechanism for the inhibition by Osx of beta-catenin transcriptional activity. We also showed that Osx decreased osteoblast proliferation. Indeed, E18.5 Osx-null calvaria showed greater BrdU incorporation than wild-type calvaria and that Osx overexpression in C2C12 mesenchymal cells inhibited cell growth. Because Wnt signaling has a major role in stimulating osteoblast proliferation, we speculate that Osx-mediated inhibition of osteoblast proliferation is a consequence of the Osx-mediated control of Wnt/beta-catenin activity. Our results add a layer of control to Wnt/beta-catenin signaling in bone.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Komori T, et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell. 1997;89:755–764. - PubMed
-
- Mundlos S, et al. Mutations involving the transcription factor CBFA1 cause cleidocranial dysplasia. Cell. 1997;89:773–779. - PubMed
-
- Nakashima K, et al. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell. 2002;108:17–29. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
