

Long-term activity-dependent plasticity of action potential propagation delay and amplitude in cortical networks
- PMID: 18461127
- PMCID: PMC2324202
- DOI: 10.1371/journal.pone.0002088
Long-term activity-dependent plasticity of action potential propagation delay and amplitude in cortical networks
Abstract
Background: The precise temporal control of neuronal action potentials is essential for regulating many brain functions. From the viewpoint of a neuron, the specific timings of afferent input from the action potentials of its synaptic partners determines whether or not and when that neuron will fire its own action potential. Tuning such input would provide a powerful mechanism to adjust neuron function and in turn, that of the brain. However, axonal plasticity of action potential timing is counter to conventional notions of stable propagation and to the dominant theories of activity-dependent plasticity focusing on synaptic efficacies.
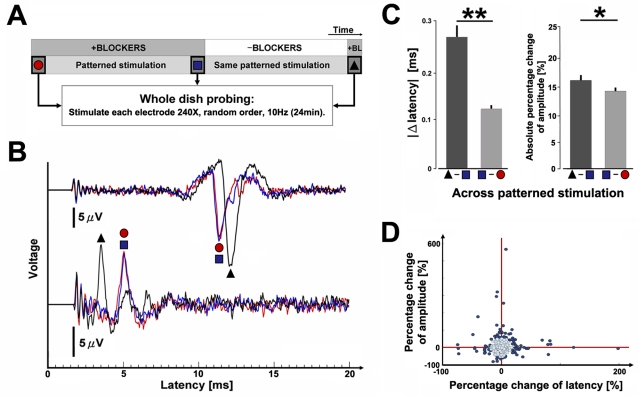
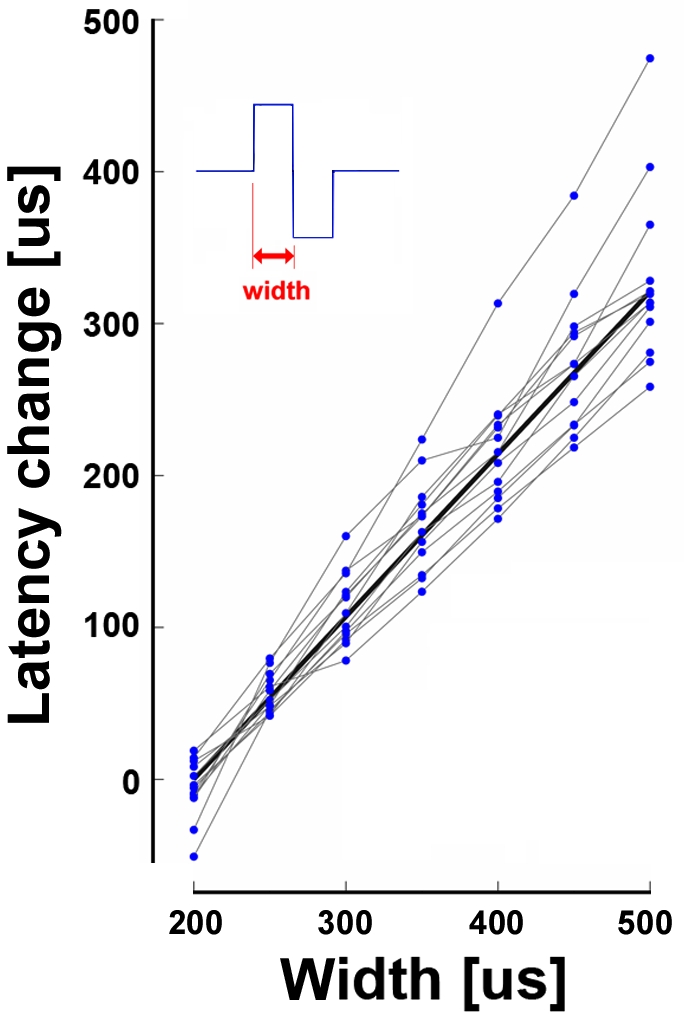
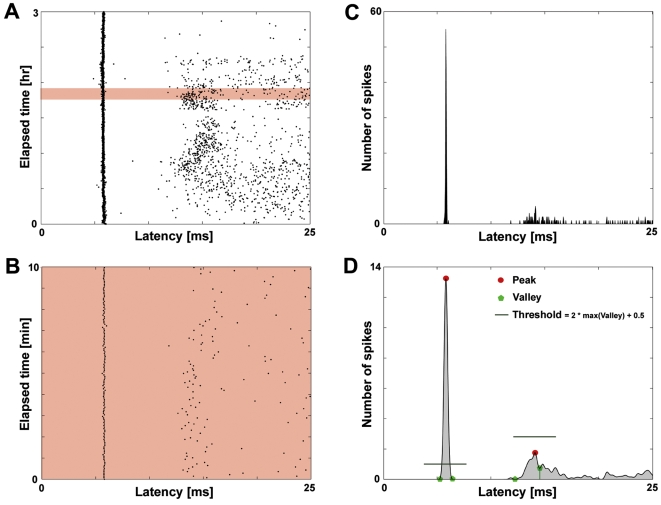
Methodology/principal findings: Here we show the occurrence of activity-dependent plasticity of action potential propagation delays (up to 4 ms or 40% after minutes and 13 ms or 74% after hours) and amplitudes (up to 87%). We used a multi-electrode array to induce, detect, and track changes in propagation in multiple neurons while they adapted to different patterned stimuli in controlled neocortical networks in vitro. The changes did not occur when the same stimulation was repeated while blocking ionotropic gabaergic and glutamatergic receptors. Even though induction of changes in action potential timing and amplitude depended on synaptic transmission, the expression of these changes persisted in the presence of the synaptic receptor blockers.
Conclusions/significance: We conclude that, along with changes in synaptic efficacy, propagation plasticity provides a cellular mechanism to tune neuronal network function in vitro and potentially learning and memory in the brain.
Conflict of interest statement
Figures
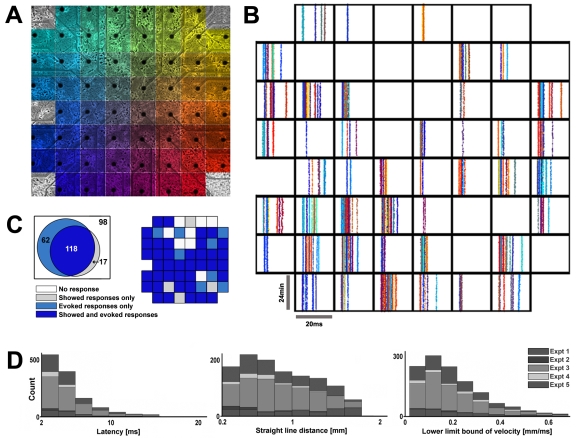
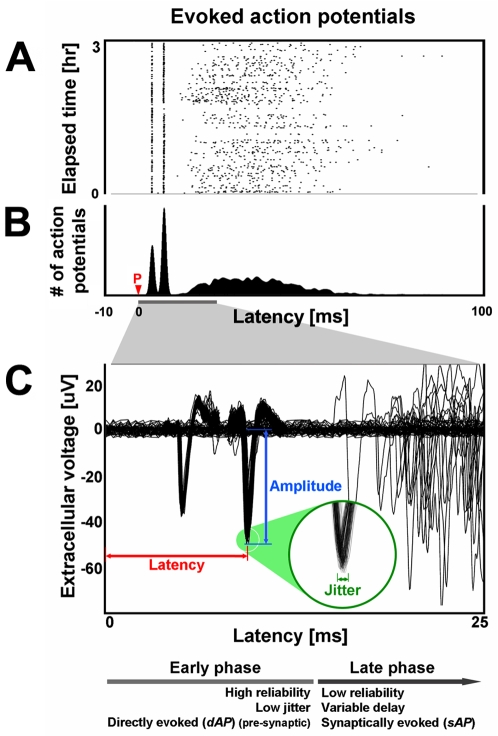
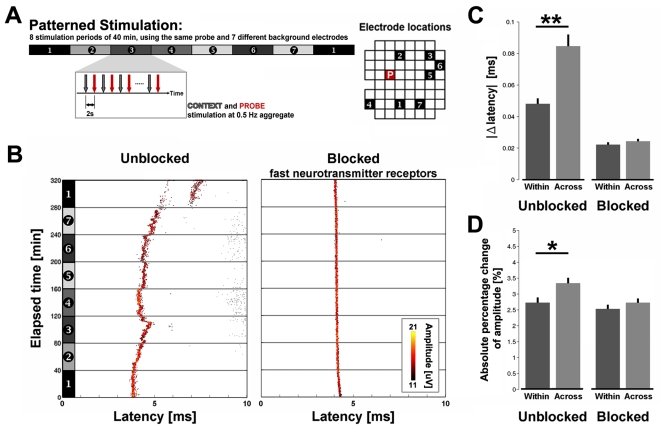
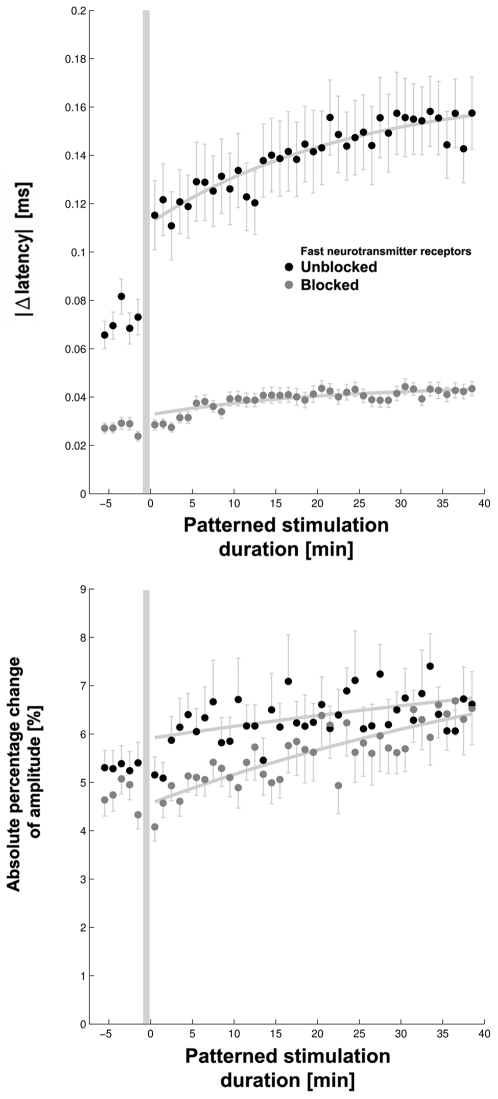
References
-
- Debanne D. Information processing in the axon. Nat Rev Neurosci. 2004;5:304–316. - PubMed
-
- Clark B, Hausser M. Neural coding: hybrid analog and digital signalling in axons. Curr Biol. 2006;16:R585–588. - PubMed
-
- Shu Y, Hasenstaub A, Duque A, Yu Y, McCormick DA. Modulation of intracortical synaptic potentials by presynaptic somatic membrane potential. Nature. 2006;441:761–765. - PubMed
-
- Alle H, Geiger JR. Combined analog and action potential coding in hippocampal mossy fibers. Science. 2006;311:1290–1293. - PubMed
-
- Izhikevich EM. Polychronization: computation with spikes. Neural Comput. 2006;18:245–282. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
