

Myosin VIIA, important for human auditory function, is necessary for Drosophila auditory organ development
- PMID: 18461180
- PMCID: PMC2362849
- DOI: 10.1371/journal.pone.0002115
Myosin VIIA, important for human auditory function, is necessary for Drosophila auditory organ development
Abstract
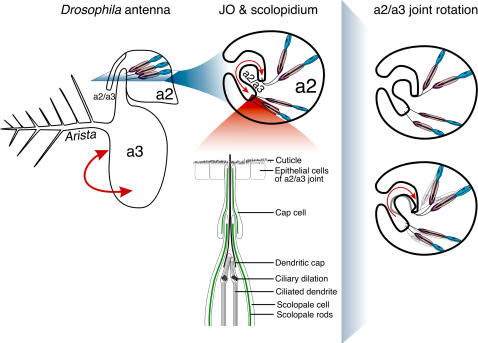
Background: Myosin VIIA (MyoVIIA) is an unconventional myosin necessary for vertebrate audition [1]-[5]. Human auditory transduction occurs in sensory hair cells with a staircase-like arrangement of apical protrusions called stereocilia. In these hair cells, MyoVIIA maintains stereocilia organization [6]. Severe mutations in the Drosophila MyoVIIA orthologue, crinkled (ck), are semi-lethal [7] and lead to deafness by disrupting antennal auditory organ (Johnston's Organ, JO) organization [8]. ck/MyoVIIA mutations result in apical detachment of auditory transduction units (scolopidia) from the cuticle that transmits antennal vibrations as mechanical stimuli to JO.
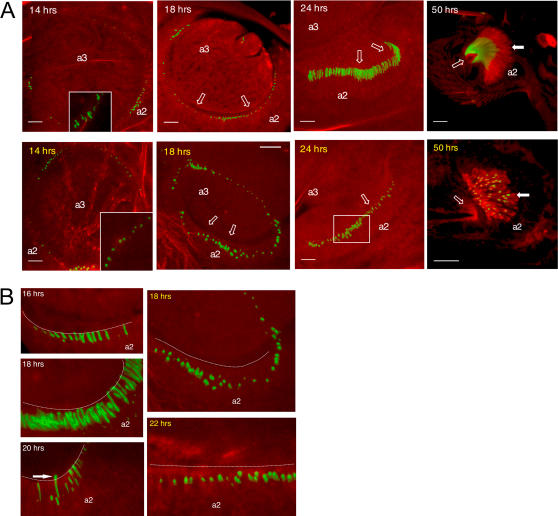
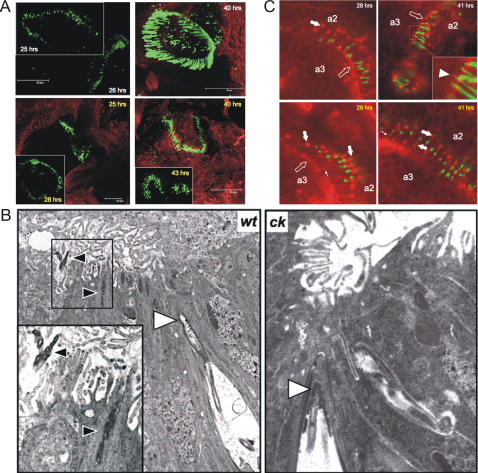
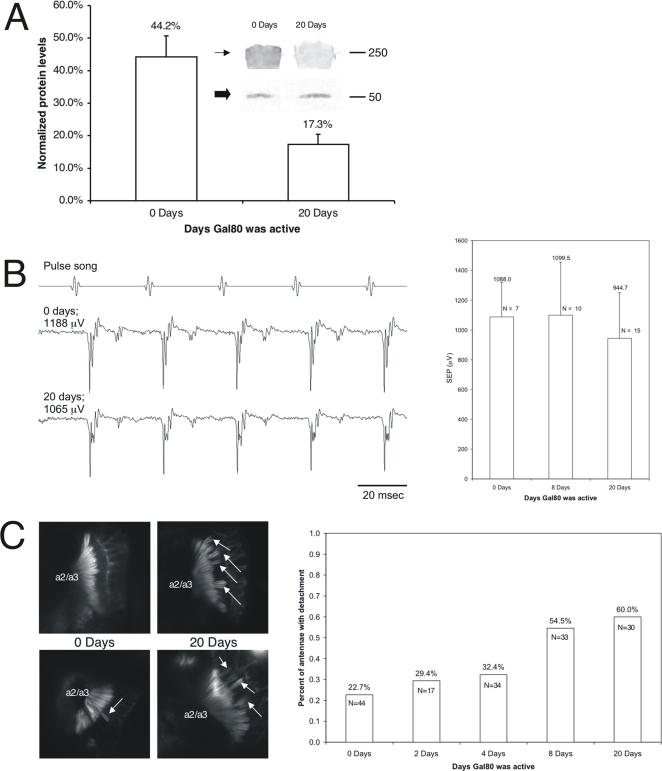
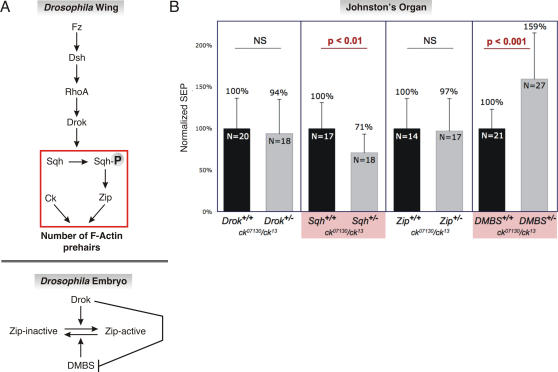
Principal findings: Using flies expressing GFP-tagged NompA, a protein required for auditory organ organization in Drosophila, we examined the role of ck/MyoVIIA in JO development and maintenance through confocal microscopy and extracellular electrophysiology. Here we show that ck/MyoVIIA is necessary early in the developing antenna for initial apical attachment of the scolopidia to the articulating joint. ck/MyoVIIA is also necessary to maintain scolopidial attachment throughout adulthood. Moreover, in the adult JO, ck/MyoVIIA genetically interacts with the non-muscle myosin II (through its regulatory light chain protein and the myosin binding subunit of myosin II phosphatase). Such genetic interactions have not previously been observed in scolopidia. These factors are therefore candidates for modulating MyoVIIA activity in vertebrates.
Conclusions: Our findings indicate that MyoVIIA plays evolutionarily conserved roles in auditory organ development and maintenance in invertebrates and vertebrates, enhancing our understanding of auditory organ development and function, as well as providing significant clues for future research.
Conflict of interest statement
Figures
References
-
- Ernest S, Rauch G-J, Haffter P, Geisler R, Petit C, et al. Mariner is defective in myosin VIIA: a zebrafish model for human hereditary deafness. Hum Mol Genet. 2000;9:2189–2196. - PubMed
-
- Gibson F, Walsh J, Mburu P, Varela A, Brown KA, et al. A type VII myosin encoded by the mouse deafness gene shaker-1. Nature. 1995;374:62–64. - PubMed
-
- Liu X-Z, Walsh J, Mburu P, Kendrick-Jones J, Cope MJTV, et al. Mutations in the myosin VIIA gene cause non-syndromic recessive deafness. Nature Genet. 1997;16:188–190. - PubMed
-
- Liu X-Z, Walsh J, Tamagawa Y, Kitamura K, Nishizawa M, et al. Autosomal dominant non-syndromic deafness caused by a mutation in the myosin VIIA gene. Nature Genet. 1997;17:268–269. - PubMed
-
- Weil D, Blanchard S, Kaplan J, Guilford P, Gibson F, et al. Defective myosin VIIA gene responsible for Usher syndrome type 1B. Nature. 1995;374:60–61. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
