

Mathematics of cell motility: have we got its number?
- PMID: 18461331
- PMCID: PMC2862828
- DOI: 10.1007/s00285-008-0182-2
Mathematics of cell motility: have we got its number?
Abstract
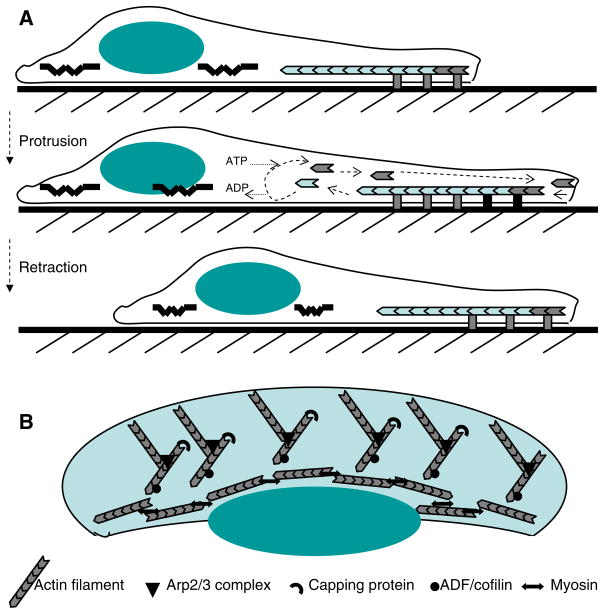
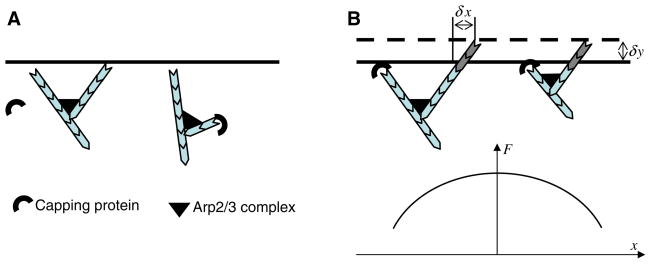
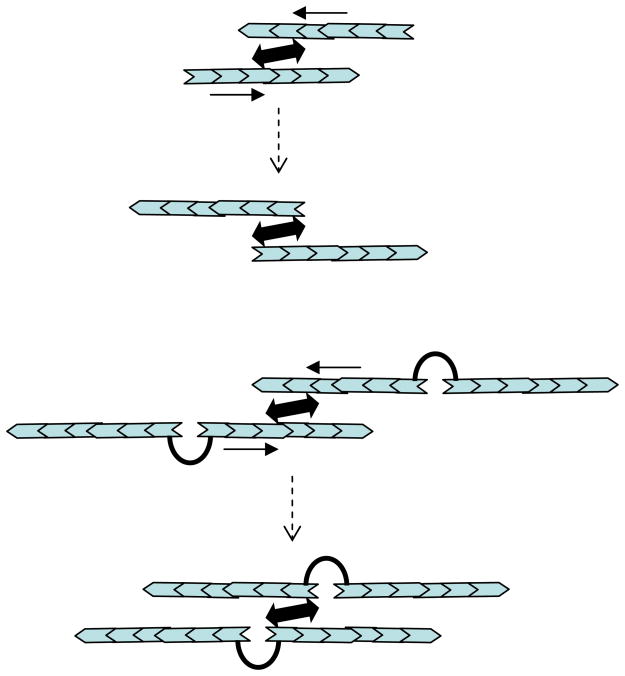
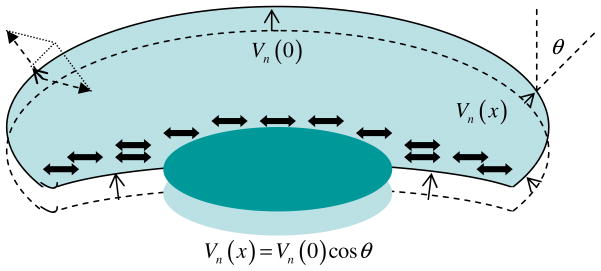
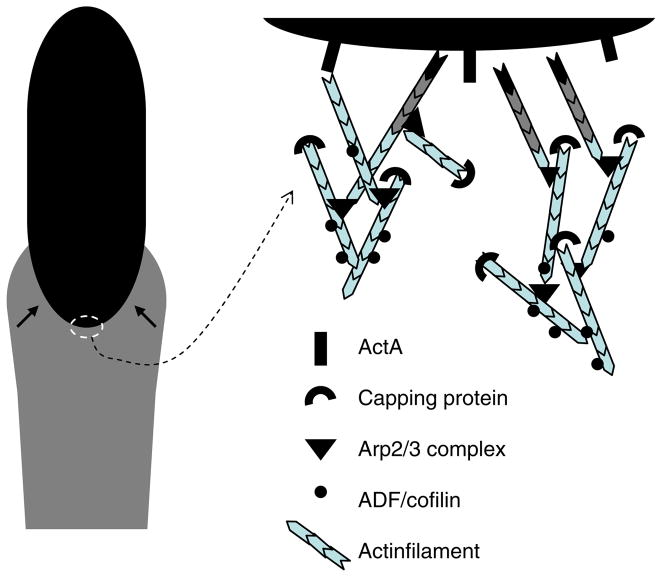
Mathematical and computational modeling is rapidly becoming an essential research technique complementing traditional experimental biological methods. However, lack of standard modeling methods, difficulties of translating biological phenomena into mathematical language, and differences in biological and mathematical mentalities continue to hinder the scientific progress. Here we focus on one area-cell motility-characterized by an unusually high modeling activity, largely due to a vast amount of quantitative, biophysical data, 'modular' character of motility, and pioneering vision of the area's experimental leaders. In this review, after brief introduction to biology of cell movements, we discuss quantitative models of actin dynamics, protrusion, adhesion, contraction, and cell shape and movement that made an impact on the process of biological discovery. We also comment on modeling approaches and open questions.
Figures





References
-
- Abercrombie M, Heaysman JE, Pegrum SM. The locomotion of fibroblasts in culture. I. Movements of the leading edge. Exp Cell Res. 1970;59:393–398. - PubMed
-
- Alt W, Dembo M. Cytoplasm dynamics and cell motion: two-phase flow models. Math Biosci. 1999;156:207–228. - PubMed
-
- Asthagiri AR, Lauffenburger DA. Bioengineering models of cell signaling. Annu Rev Biomed Eng. 2000;2:31–53. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
