

The involvement of the tyrosine kinase c-Src in the regulation of reactive oxygen species generation mediated by NADPH oxidase-1
- PMID: 18463161
- PMCID: PMC2441655
- DOI: 10.1091/mbc.e08-02-0138
The involvement of the tyrosine kinase c-Src in the regulation of reactive oxygen species generation mediated by NADPH oxidase-1
Abstract
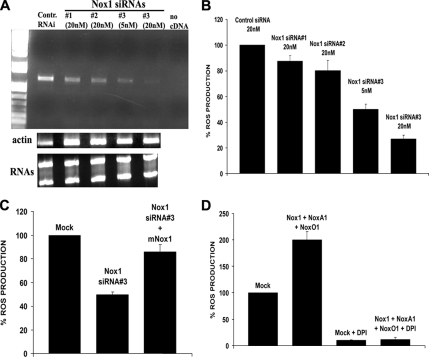
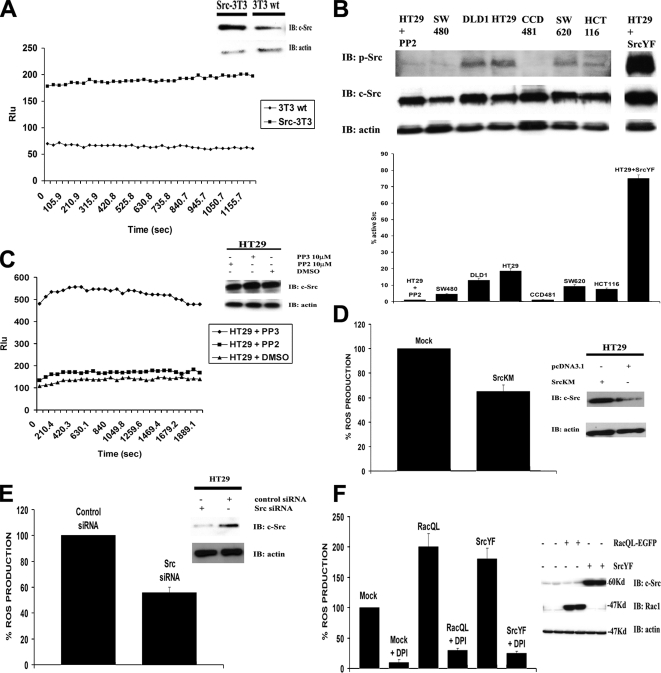
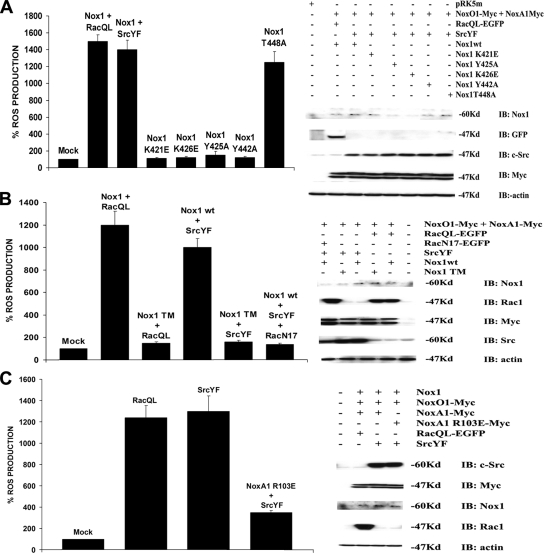
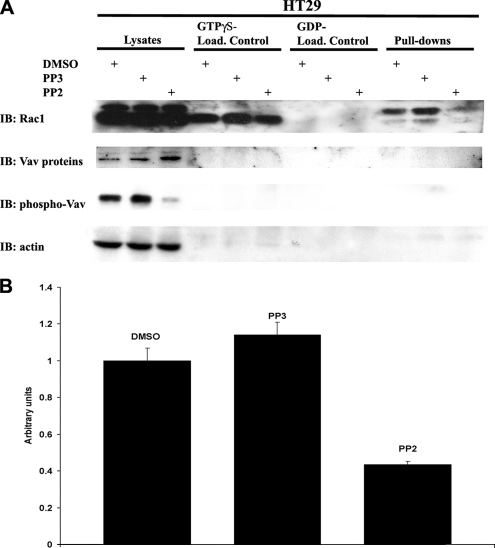
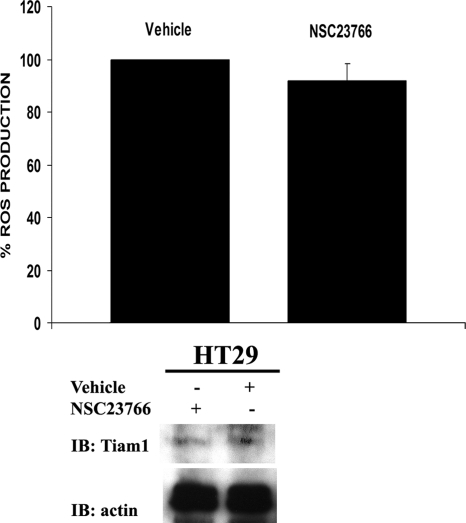
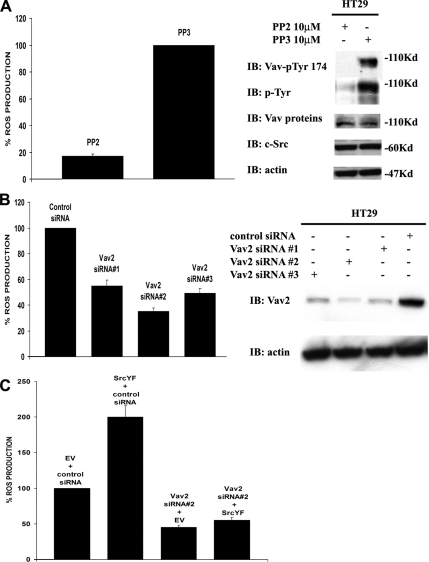
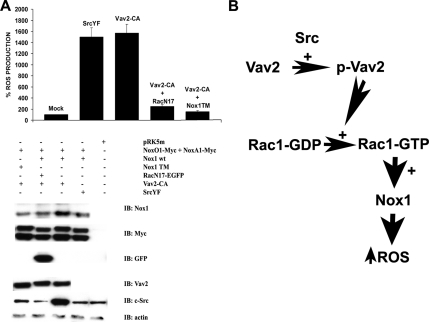
NADPH oxidase (Nox) family enzymes are one of the main sources of cellular reactive oxygen species (ROS), which have been shown to function as second messenger molecules. To date, seven members of this family have been reported, including Nox1-5 and Duox1 and -2. With the exception of Nox2, the regulation of the Nox enzymes is still poorly understood. Nox1 is highly expressed in the colon, and it requires two cytosolic regulators, NoxO1 and NoxA1, as well as the binding of Rac1 GTPase, for its activity. In this study, we investigate the role of the tyrosine kinase c-Src in the regulation of ROS formation by Nox1. We show that c-Src induces Nox1-mediated ROS generation in the HT29 human colon carcinoma cell line through a Rac-dependent mechanism. Treatment of HT29 cells with the Src inhibitor PP2, expression of a kinase-inactive form of c-Src, and c-Src depletion by small interfering RNA (siRNA) reduce both ROS generation and the levels of active Rac1. This is associated with decreased Src-mediated phosphorylation and activation of the Rac1-guanine nucleotide exchange factor Vav2. Consistent with this, Vav2 siRNA that specifically reduces endogenous Vav2 protein is able to dramatically decrease Nox1-dependent ROS generation and abolish c-Src-induced Nox1 activity. Together, these results establish c-Src as an important regulator of Nox1 activity, and they may provide insight into the mechanisms of tumor formation in colon cancers.
Figures
References
-
- Abe J., Takahashi M., Ishida M., Lee J. D., Berk B. C. c-Src is required for oxidative stress-mediated activation of big mitogen-activated protein kinase 1. J. Biol. Chem. 1997;272:20389–20394. - PubMed
-
- Abid M. R., Kachra Z., Spokes K. C., Aird W. C. NADPH oxidase activity is required for endothelial cell proliferation and migration. FEBS Lett. 2000;486:252–256. - PubMed
-
- Abram C. L., Courtneidge S. A. Src family tyrosine kinases and growth factor signaling. Exp. Cell Res. 2000;254:1–13. - PubMed
-
- Alvarez R. H., Kantarjian H. M., Cortes J. E. The role of Src in solid and hematologic malignancies: development of new-generation Src inhibitors. Cancer. 2006;107:1918–1929. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
