

Learning to recognize visual objects with microstimulation in inferior temporal cortex
- PMID: 18463185
- PMCID: PMC2493485
- DOI: 10.1152/jn.90247.2008
Learning to recognize visual objects with microstimulation in inferior temporal cortex
Abstract
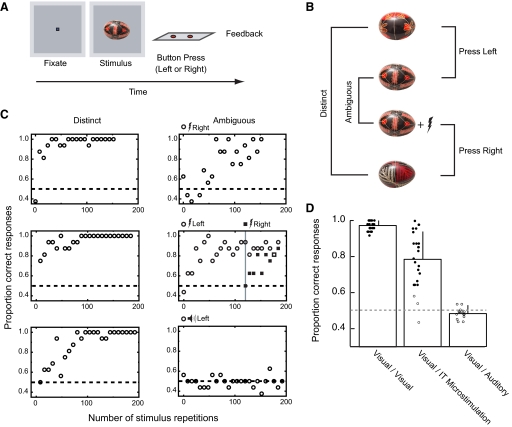
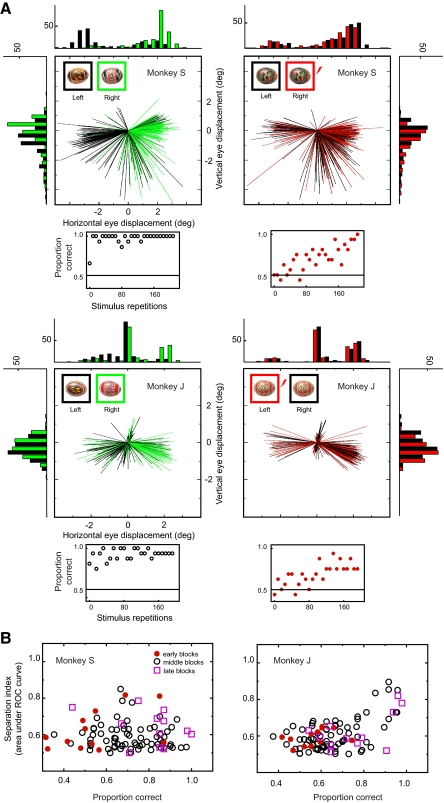
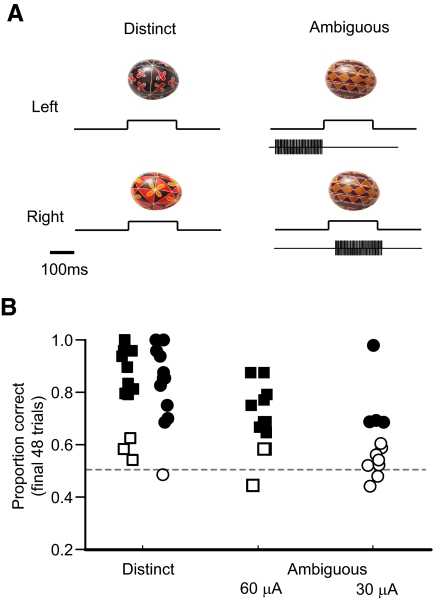
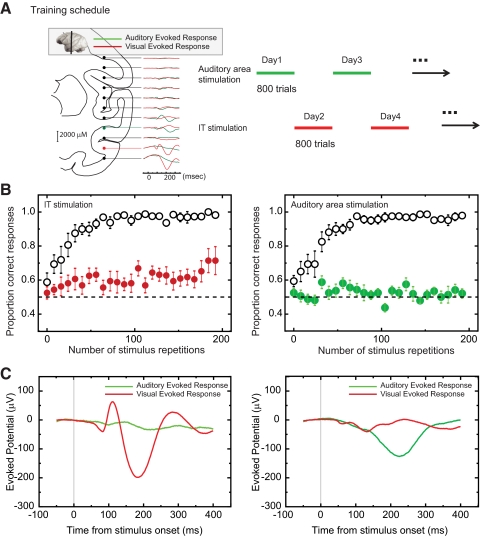
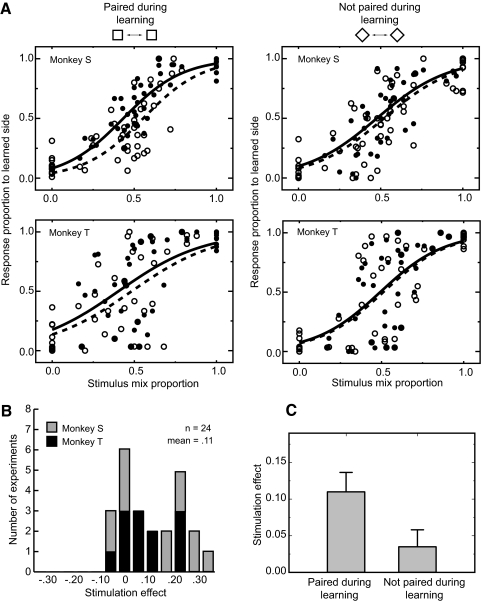
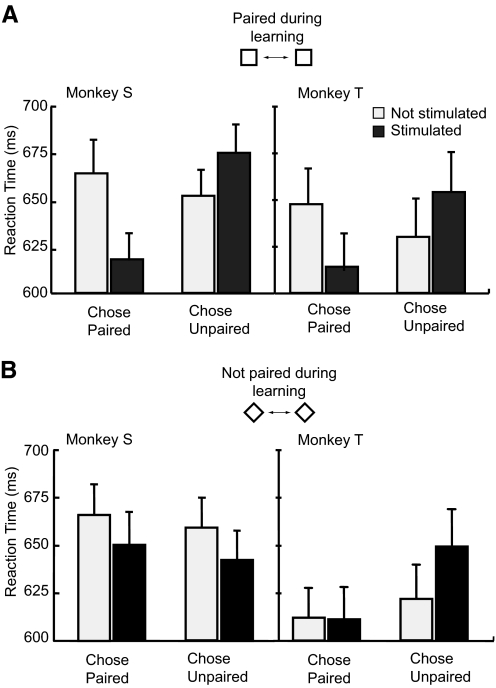
The malleability of object representations by experience is essential for adaptive behavior. It has been hypothesized that neurons in inferior temporal cortex (IT) in monkeys are pivotal in visual association learning, evidenced by experiments revealing changes in neural selectivity following visual learning, as well as by lesion studies, wherein functional inactivation of IT impairs learning. A critical question remaining to be answered is whether IT neuronal activity is sufficient for learning. To address this question directly, we conducted experiments combining visual classification learning with microstimulation in IT. We assessed the effects of IT microstimulation during learning in cases where the stimulation was exclusively informative, conditionally informative, and informative but not necessary for the classification task. The results show that localized microstimulation in IT can be used to establish visual classification learning, and the same stimulation applied during learning can predictably bias judgments on subsequent recognition. The effect of induced activity can be explained neither by direct stimulation-motor association nor by simple detection of cortical stimulation. We also found that the learning effects are specific to IT stimulation as they are not observed by microstimulation in an adjacent auditory area. Our results add the evidence that the differential activity in IT during visual association learning is sufficient for establishing new associations. The results suggest that experimentally manipulated activity patterns within IT can be effectively combined with ongoing visually induced activity during the formation of new associations.
Figures


References
-
- Afraz SR, Kiani R, Esteky H. Microstimulation of inferotemporal cortex influences face categorization. Nature 442: 692–695, 2006. - PubMed
-
- Baker CI, Behrmann M, Olson CR. Impact of learning on representation of parts and wholes in monkey inferotemporal cortex. Nat Neurosci 5: 1210–1216, 2002. - PubMed
-
- Bisley JW, Zaksas D, Pasternak T. Microstimulation of cortical area MT affects performance on a visual working memory task. J Neurophysiol 85: 187–196, 2001. - PubMed
-
- Bolster B, Crowne DP. Effects of anterior and posterior inferotemporal lesions on discrimination reversal in the monkey. Neuropsychologia 17: 11–20, 1979. - PubMed
-
- Bradley DC, Troyk PR, Berg JA, Bak M, Cogan S, Erickson R, Kufta C, Mascaro M, McCreery D, Schmidt EM, Towle VL, Xu H. Visuotopic mapping through a multichannel stimulating implant in primate V1. J Neurophysiol 93: 1659–1670, 2005. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources