

Inflammation-induced shift in the valence of spinal GABA-A receptor-mediated modulation of nociception in the adult rat
- PMID: 18467182
- PMCID: PMC2581496
- DOI: 10.1016/j.jpain.2008.03.004
Inflammation-induced shift in the valence of spinal GABA-A receptor-mediated modulation of nociception in the adult rat
Abstract
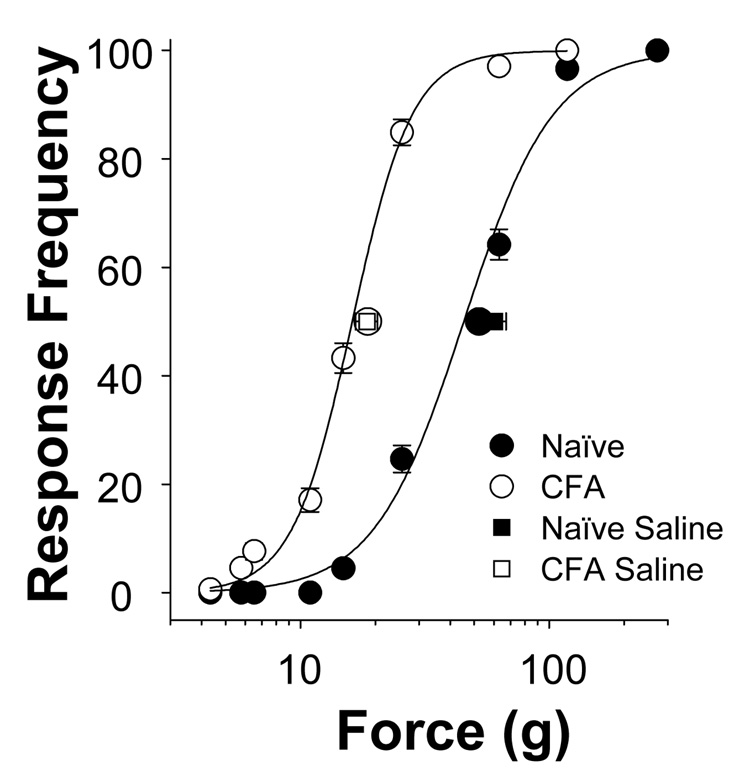
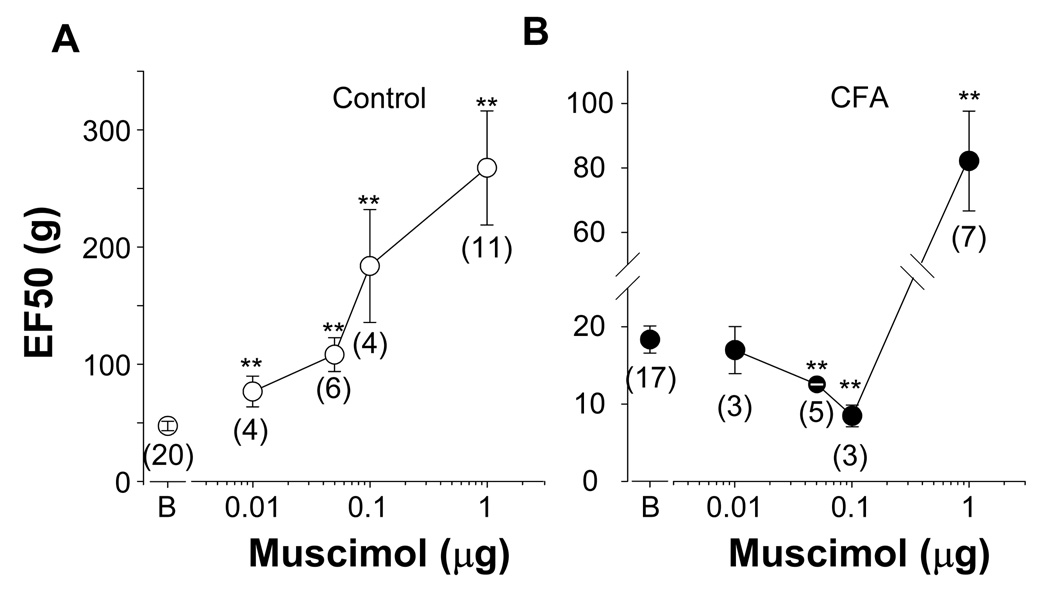
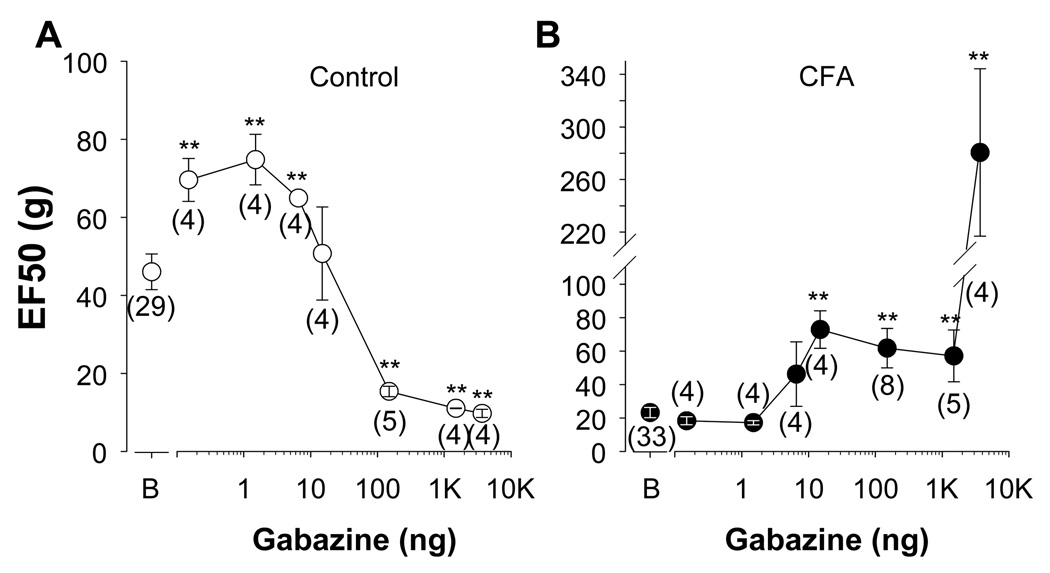
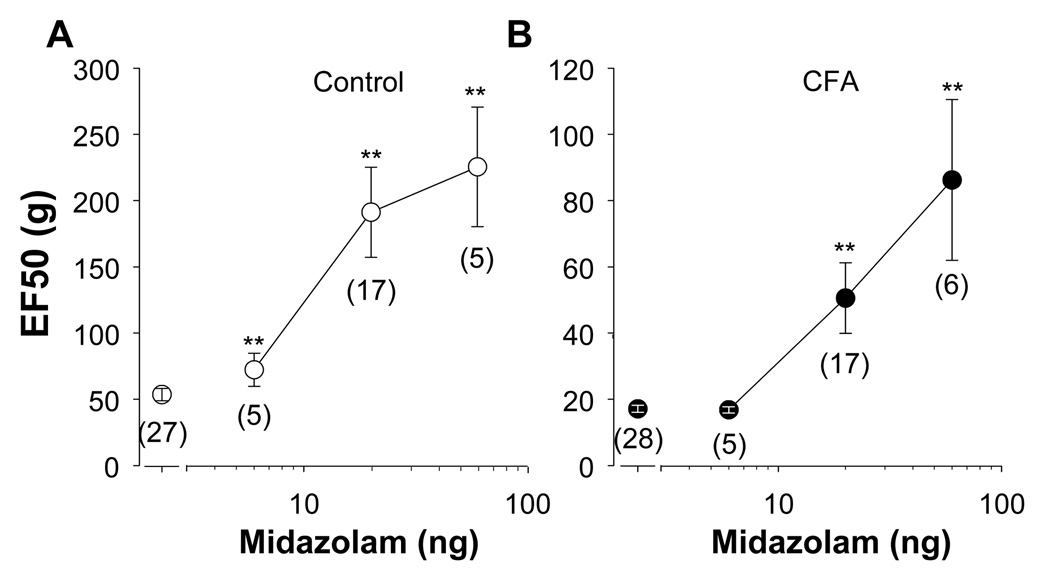
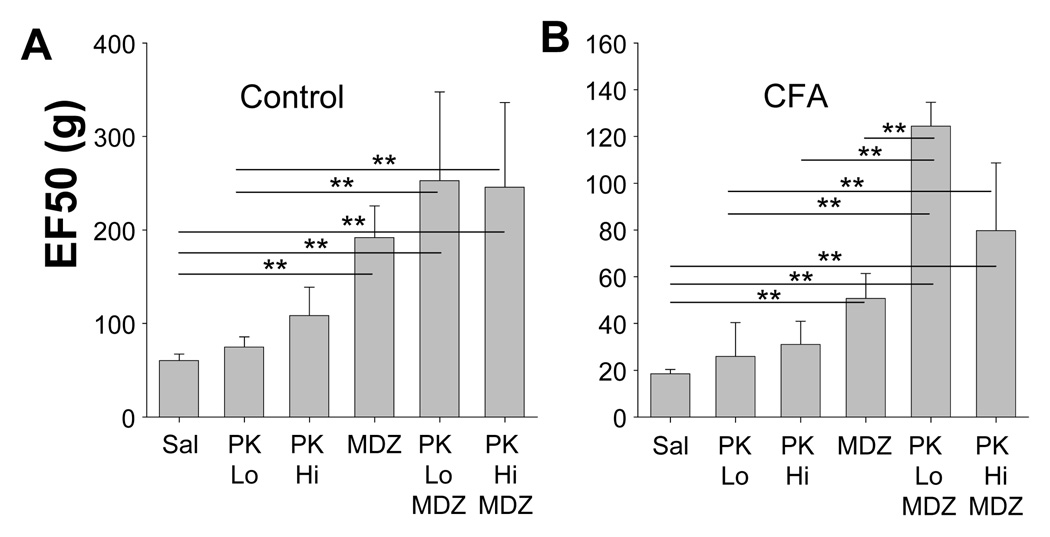
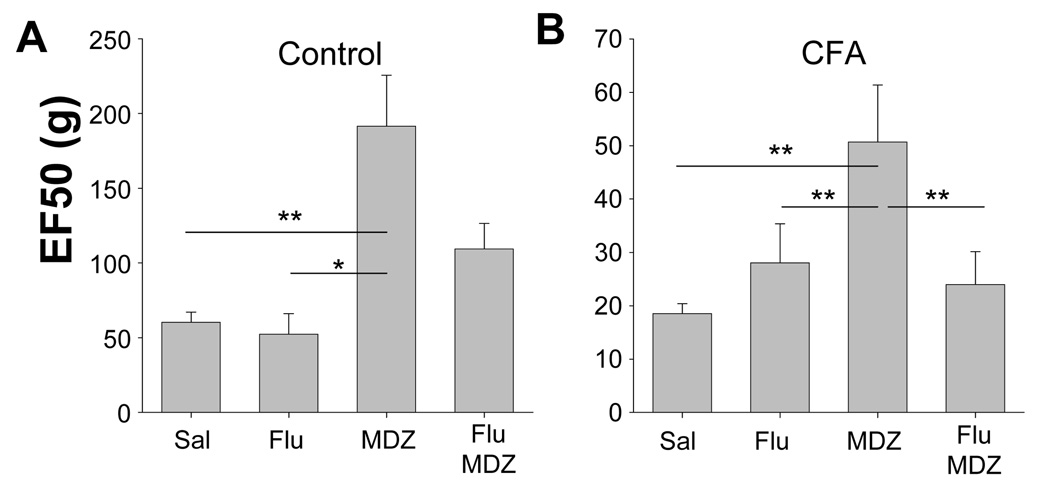
The objective of this study was to assess the impact of persistent inflammation on spinal gamma-aminobutyric acid-A (GABA-A) receptor-mediated modulation of evoked nociceptive behavior in the adult rat. Nocifensive threshold was assessed with von Frey filaments applied to the dorsal surface of the hind paw. The GABA-A receptor agonist muscimol, the antagonist gabazine, the benzodiazepine receptor agonist midazolam, and antagonists PK11195 and flumazenil were administered spinally in the presence and absence of complete Freund's adjuvant (CFA)-induced inflammation. In naive rats, muscimol increased and gabazine decreased nociceptive threshold. After CFA, the effects of these compounds were reversed: Low doses of muscimol exacerbated the inflammation-induced decrease in nociceptive threshold and gabazine increased nociceptive threshold. Midazolam increased nociceptive threshold both in the presence and absence of inflammation. Flumazenil but not PK11195 blocked the analgesic effects of midazolam. These findings indicate that inflammation-induced changes in GABA-A signaling are complex and are likely to involve several distinct mechanisms. Rectifying the changes in GABA-A signaling may provide effective relief from hypersensitivity observed in the presence of inflammation.
Perspective: An inflammation-induced shift in spinal GABA-A receptor signaling from inhibition to excitation appears to underlie inflammatory pain and hypersensitivity. Use of GABA-A receptor selective general anesthetics in association with therapeutic interventions may be contraindicated. More importantly, rectifying the changes in GABA-A signaling may provide effective relief from inflammatory hypersensitivity.
Figures
References
-
- Baba H, Ji RR, Kohno T, Moore KA, Ataka T, Wakai A, Okamoto M, Woolf CJ. Removal of GABAergic inhibition facilitates polysynaptic A fiber-mediated excitatory transmission to the superficial spinal dorsal horn. Mol Cell Neurosci. 2003;24:818–830. - PubMed
-
- Barber RP, Vaughn JE, Saito K, McLaughlin BJ, Roberts E. GABAergic terminals are presynaptic to primary afferent terminals in the substantia gelatinosa of the rat spinal cord. Brain Res. 1978;141:35–55. - PubMed
-
- Brady LS, Mansbach RS, Skurdal DN, Muldoon SM, Barrett JE. Reversal of the antinociceptive effects of centrally-administered morphine by the benzodiazepine receptor antagonist Ro 15-1788. Life Sci. 1984;35:2593–2600. - PubMed
-
- Buhr A, Baur R, Sigel E. Subtle changes in residue 77 of the gamma subunit of alpha1beta2gamma2 GABAA receptors drastically alter the affinity for ligands of the benzodiazepine binding site. J Biol Chem. 1997;272:11799–11804. - PubMed
-
- Castro-Lopes JM, Tolle TR, Pan B, Zieglgansberger W. Expression of GAD mRNA in spinal cord neurons of normal and monoarthritic rats. Brain Res Mol Brain Res. 1994;26:169–176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
