

The androgen receptor is selectively involved in organization of sexually dimorphic social behaviors in mice
- PMID: 18467440
- PMCID: PMC2488208
- DOI: 10.1210/en.2008-0183
The androgen receptor is selectively involved in organization of sexually dimorphic social behaviors in mice
Abstract
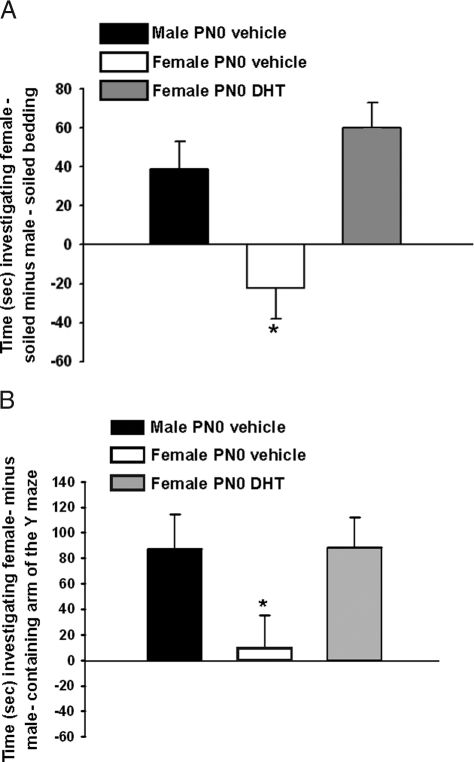
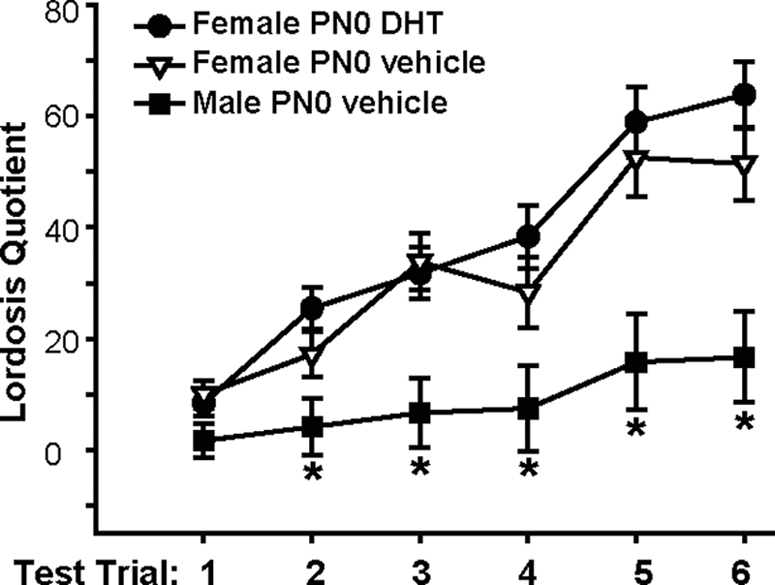
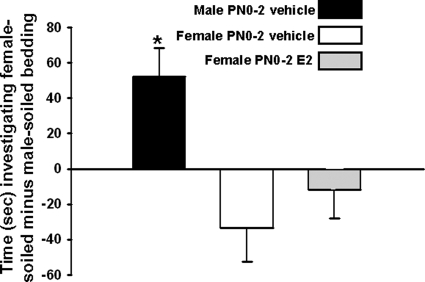
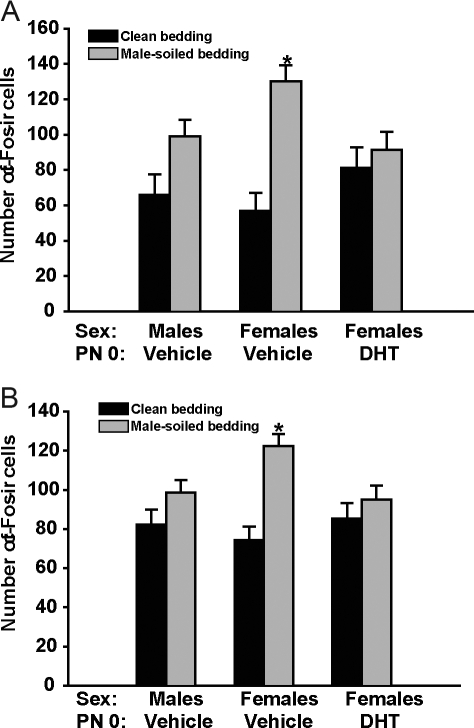
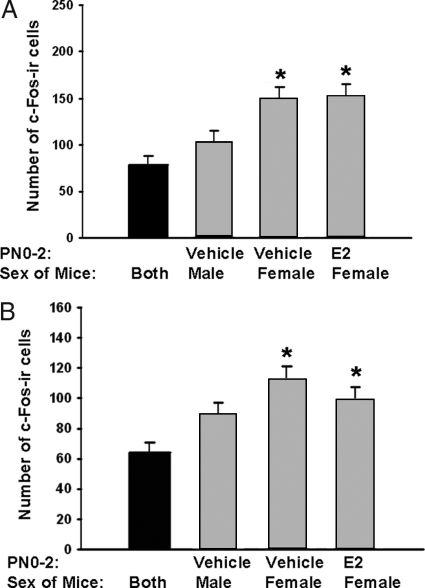
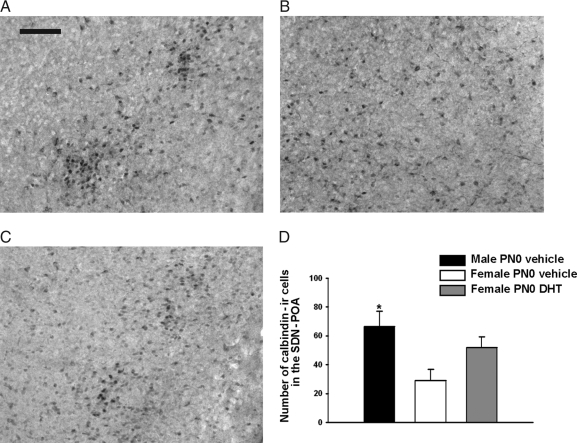
It is well established that sexually dimorphic neural regions are organized by steroid hormones during development. In many species, neonatal males are exposed to more testosterone than their female littermates, and ultimately it is the estradiol, produced by aromatization of testosterone, that affects sexual differentiation. However, the androgen receptor also plays an important role in the masculinization of brain and behavior. Here we tested the hypothesis that sexually dimorphic social and odor preference behaviors can be differentiated by a nonaromatizable androgen during development by treating female mice on the day of birth (PN0) with dihydrotestosterone (DHT). Control mice received a single vehicle injection on PN0. Adults were gonadectomized, treated with estradiol, and tested for social behaviors. In contrast with control females, females treated on PN0 with DHT, like male controls, exhibited a preference for female-soiled vs. male-soiled bedding, a preference to investigate a female vs. a male and reduced c-Fos-immunoreactivity (ir) in several neural areas after exposure to male-soiled bedding. However, females treated with DHT on PN0 had normal female-typical sexual behavior. The number of calbindin-ir cells in the preoptic area is sexually dimorphic (males more than females), but females given DHT on PN0 had intermediate numbers of calbindin-ir neurons, not significantly different from control males or females. Our data demonstrate that organization of social and olfactory preferences in mice can be affected by perinatal DHT and lends support to the role of androgen receptor in organization of sexual differentiation of brain and behaviors.
Figures
Similar articles
-
Androgen receptor is essential for sexual differentiation of responses to olfactory cues in mice.Eur J Neurosci. 2007 Apr;25(7):2182-90. doi: 10.1111/j.1460-9568.2007.05484.x. Eur J Neurosci. 2007. PMID: 17419752
-
Non-neural androgen receptor promotes androphilic odor preference in mice.Horm Behav. 2016 Jul;83:14-22. doi: 10.1016/j.yhbeh.2016.05.013. Epub 2016 May 15. Horm Behav. 2016. PMID: 27191855
-
Prenatal influence of an androgen agonist and antagonist on the differentiation of the ovine sexually dimorphic nucleus in male and female lamb fetuses.Endocrinology. 2014 Dec;155(12):5000-10. doi: 10.1210/en.2013-2176. Epub 2014 Sep 12. Endocrinology. 2014. PMID: 25216387 Free PMC article.
-
Steroid-induced plasticity in the sexually dimorphic vasotocinergic innervation of the avian brain: behavioral implications.Brain Res Brain Res Rev. 2001 Nov;37(1-3):178-200. doi: 10.1016/s0165-0173(01)00118-7. Brain Res Brain Res Rev. 2001. PMID: 11744086 Review.
-
Hormonal influences on sexually differentiated behavior in nonhuman primates.Front Neuroendocrinol. 2005 Apr;26(1):7-26. doi: 10.1016/j.yfrne.2005.02.001. Front Neuroendocrinol. 2005. PMID: 15862182 Review.
Cited by
-
Role of neural stem cell activity in postweaning development of the sexually dimorphic nucleus of the preoptic area in rats.PLoS One. 2013;8(1):e54927. doi: 10.1371/journal.pone.0054927. Epub 2013 Jan 30. PLoS One. 2013. PMID: 23383001 Free PMC article.
-
Potential contribution of prenatal estrogens to the sexual differentiation of mate preferences in mice.Horm Behav. 2011 Jan;59(1):83-9. doi: 10.1016/j.yhbeh.2010.10.012. Epub 2010 Oct 26. Horm Behav. 2011. PMID: 21029737 Free PMC article.
-
Calbindin Knockout Alters Sex-Specific Regulation of Behavior and Gene Expression in Amygdala and Prefrontal Cortex.Endocrinology. 2016 May;157(5):1967-79. doi: 10.1210/en.2016-1055. Epub 2016 Mar 24. Endocrinology. 2016. PMID: 27010449 Free PMC article.
-
Neonatal testosterone partially organizes sex differences in stress-induced emotionality in mice.Neurobiol Dis. 2012 May;46(2):486-96. doi: 10.1016/j.nbd.2012.02.014. Epub 2012 Feb 28. Neurobiol Dis. 2012. PMID: 22394611 Free PMC article.
-
Moxd1 Is a Marker for Sexual Dimorphism in the Medial Preoptic Area, Bed Nucleus of the Stria Terminalis and Medial Amygdala.Front Neuroanat. 2017 Mar 27;11:26. doi: 10.3389/fnana.2017.00026. eCollection 2017. Front Neuroanat. 2017. PMID: 28396628 Free PMC article.
References
-
- Phoenix CH, Goy RW, Gerall AA, Young WC 1959 Organizing action of prenatally administered testosterone propionate on the tissues mediating mating behavior in the female guinea pig. Endocrinology 65:369–382 - PubMed
-
- Negri-Cesi P, Colciago A, Celotti F, Motta M 2004 Sexual differentiation of the brain: role of testosterone and its active metabolites. J Endocrinol Invest 27:120–127 - PubMed
-
- Corbier P, Edwards DA, Roffi J 1992 The neonatal testosterone surge: a comparative study. Arch Int Physiol Biochim Biophys 100:127–131 - PubMed
-
- Vomachka AJ, Lisk RD 1986 Androgen and estradiol levels in plasma and amniotic fluid of late gestational male and female hamsters: uterine position effects. Horm Behav 20:181–193 - PubMed
-
- Vreeburg JT, Groeneveld JO, Post PE, Ooms MP 1983 Concentrations of testosterone and androsterone in peripheral and umbilical venous plasma of fetal rats. J Reprod Fertil 68:171–175 - PubMed
