

T-cell factor 3 regulates embryonic stem cell pluripotency and self-renewal by the transcriptional control of multiple lineage pathways
- PMID: 18467660
- PMCID: PMC2692055
- DOI: 10.1634/stemcells.2007-1115
T-cell factor 3 regulates embryonic stem cell pluripotency and self-renewal by the transcriptional control of multiple lineage pathways
Abstract
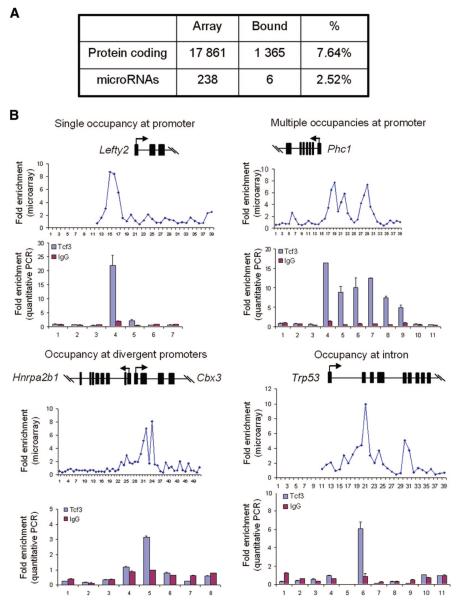
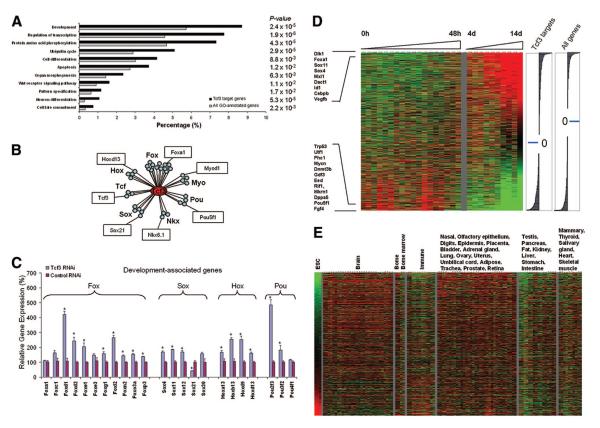
The Wnt signaling pathway is necessary both for maintaining undifferentiated stem cells and for directing their differentiation. In mouse embryonic stem cells (ESCs), Wnt signaling preferentially maintains "stemness" under certain permissive conditions. T-cell factor 3 (Tcf3) is a component of the Wnt signaling and a dominant downstream effector in ESCs. Despite the wealth of knowledge regarding the importance of Wnt signaling underlying stem cells functions, the precise mechanistic explanation by which the effects are mediated is unknown. In this study, we identified new regulatory targets of Tcf3 using a whole-genome approach and found that Tcf3 transcriptionally represses many genes important for maintaining pluripotency and self-renewal, as well as those involved in lineage commitment and stem cell differentiation. This effect is in part mediated by the corepressors transducin-like enhancer of split 2 and C-terminal Binding Protein (CtBP). Notably, Tcf3 binds to and represses the Oct4 promoter, and this repressive effect requires both the Groucho and CtBP interacting domains of Tcf3. Interestingly, we find that in mouse preimplantation development embryos, Tcf3 expression is coregulated with Oct4 and Nanog and becomes localized to the inner cell mass of the blastocyst. These data demonstrate an important role for Tcf3 in modulating the appropriate level of gene transcription in ESCs and during embryonic development. Disclosure of potential conflicts of interest is found at the end of this article.
Figures





References
-
- Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol. 2004;20:781–810. - PubMed
-
- Zechner D, Fujita Y, Hulsken J, et al. beta-Catenin signals regulate cell growth and the balance between progenitor cell expansion and differentiation in the nervous system. Dev Biol. 2003;258:406–418. - PubMed
-
- Hirabayashi Y, Itoh Y, Tabata H, et al. The Wnt/beta-catenin pathway directs neuronal differentiation of cortical neural precursor cells. Development. 2004;131:2791–2801. - PubMed
-
- Huelsken J, Vogel R, Erdmann B, et al. beta-Catenin controls hair follicle morphogenesis and stem cell differentiation in the skin. Cell. 2001;105:533–545. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
